











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
La familia Poaceae comprende 11783 especies, 789 géneros y 12 subfamilias; presenta una distribución cosmopolita en todos los continentes (Soreng et al. 2022). En Perú Hitchcock (1927) aporta el primer tratamiento acerca de los taxones, distribución y otros aspectos de las gramíneas junto a los de Ecuador y Bolivia; posteriormente Standley (1936) en la flora de Perú, reporta 400 especies incluidas en 102 géneros. Varias décadas después se presenta una de las mayores contribuciones con el tratamiento taxonómico de Tovar (1993), quien da a conocer 680 especies y 152 géneros; y en el mismo año se presenta el catálogo de Brako y Zarucchi (1993) donde reconocen 719 especies y 150 géneros; Ulloa Ulloa et al. (2004) adicionan 32 taxones hasta ese año.
En los últimos 20 años el conocimiento de la flora agrostológica para Perú se ha incrementado con lo mencionado en los catálogos de gramíneas del nuevo mundo (Soreng et al. 2003, Zuloaga et al. 2003, Judziewicz et al. 2014, Peterson et al. 2015); tratamientos taxonómicos en varios géneros, tales como: Aristida, Bromus, Cenchrus, Chusquea, Eragrostis, Muhlenbergia, Poa, Sporobolus, Trichachne y otros (Cialdella & Arriaga 1998, Cialdella & Giussani 2002, Saarela et al. 2006, Peterson & Sánchez-Vega 2007, Giraldo-Cañas & Peterson 2009, Giraldo-Cañas et al. 2012, Guerreiro et al. 2014, Gutiérrez 2015, Sylvester et al. 2016, Alegría et al. 2017, Lo Medico et al. 2017, Peterson et al. 2018, Fadrique et al. 2019, Gutiérrez et al. 2018, 2019, 2021). Así mismo, la ampliación del conocimiento de las gramíneas se dio mediante contribuciones locales como: en el Parque Nacional Yanachaga-Chemillén (La Torre et al. 2003, 2004), la costa norte de Perú (Tovar 2005), el río Los Amigos de Madre de Dios (Olivier 2008), Loreto (Tovar 2009), el distrito de Arahuay en Lima (Gonzáles et al. 2011) y Huancavelica (Gutiérrez & Castañeda 2014, 2017).
Poaceae en las formaciones de lomas costera de Perú, con 43 géneros y 96 especies, es la tercera familia con mayor representación después de Solanaceae y Asteraceae (Song et al. 2023). En la última década, el número de géneros se incrementó en 11.5% y en 12.5% el de especies (Dillon et al. 2011). Del total de especies de gramíneas de las formaciones de lomas cerca del 70% son nativas y 3% son endémicas (Dillon et al. 2011). Además, los estudios florísticos a lo largo de la costa peruana permiten delimitar los patrones de riqueza y distribución de las gramíneas (Cano et al. 1999, Arakaki & Cano 2003, Leiva et al. 2008, 2014, Quipuscoa et al. 2016, Talavera et al. 2017, Cuba-Melly & Odar 2018).
El objetivo del presente estudio es el dar a conocer dos nuevos registros para la flora agrostológica de Perú; así mismo, se discuten las diferencias morfológicas con taxones afines y se brindan datos ecológicos sobre ambos taxones.
Material y métodos
Durante los meses de noviembre y diciembre en los años 2020 y 2021 se realizaron expediciones a las formaciones de lomas del departamento de Arequipa, principalmente para la recolección y obtención de datos de las gramíneas; los taxones reportados en el presente estudio fueron colectados en la provincia de Caravelí (Figura 1). Para la determinación de los especímenes se utilizó claves o revisiones taxonómicas especializadas, para Piptochetium (Cialdella & Arriaga 1998, Cialdella & Giussani 2002) y Trichachne (Rúgolo de Agrasar 1974, Vega & Rúgolo de Agrasar 2002, 2005, Giraldo-Cañas 2005, Lo Medico et al. 2017, Sánchez-Ken 2012). Se contrastó con los protólogos y la serie de tipos digitalizados en el sitio web de JSTOR Global Plant (JSTOR 2023). Para cada taxón se presenta: una clasificación supraespecífica según Soreng et al. (2022), descripción morfológica, fotografías, ilustraciones, mapa de distribución y clave taxonómica para los taxones afines. Para la elaboración de las claves taxonómicas se consideró: las descripciones e información de Cialdella y Arriaga (1998) para aquellas especies con el antecio obconico de la sección Podopogon (Raf.) Parodi de Piptochetium, además, Rúgolo de Agrasar (1974), Sánchez-Ken (2012) y Lo Medico et al. (2017) para los taxones de Trichachne con espiguillas de indumento blanquecino-plateado o blanquecino-violáceo y la lemma inferior con tres nervios expuestos en el dorso.
Taxonomía
Familia: Poaceae Barnhart
Subfamilia: Pooideae Benth.
Supertribu: Stipodae L. Liu
Tribu: Stipeae Dumort.
Género: Piptochaetium J. Presl
Piptochaetium bicolor (Vahl) É. Desv.
Figuras 1, 2A-D y 3A-E
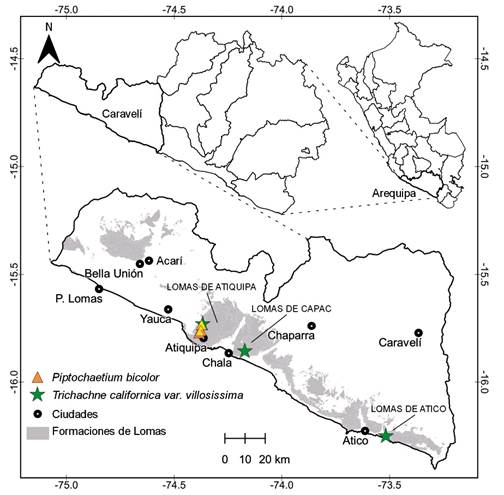
Figura 1 Distribución de Piptochaetium bicolor y Trichachne californica var. villosissima. La delimitación de las formaciones de lomas está basada en Moat et al. 2021.

Figura 2 A-D. Piptochaetium bicolor. A. Hábitat (Lomas de Atiquipa). B. Hábito. C. Panícula. D. Antecio. E-H. Trichachne californica var. villosissima. E. Hábitat (Lomas de Capac). F. Hábito. G. Panícula. H. Espiguilla. Fotografías de J. Muñuico.
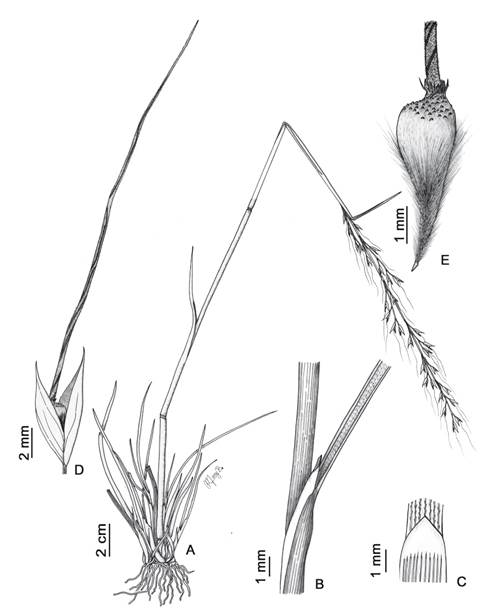
Figura 3 Piptochaetium bicolor. A. Hábito. B. Área ligular de la hoja. C. lígula. D. Espiguilla. E. Antecio. Ilustrado por Maricruz Bedoya, a partir de J. Muñuico & M. Bedoya 802 (HSP).
Fl. Chil. 6:273 (1854).
=Stipa bicolor Vahl, Symb. Bot. 2: 24 (1791).
=Oryzopsis bicolor (Vahl) Speg., Anales Mus. Nac. Montevideo 4(2): 6 (1901).
Tipo. Uruguay, "Habitat in Brasiliae, Monte Video”, Dn. Thouin s. n. (holotipo C!).
Hierba perenne, de 20-80 cm de alto. Culmos erguidos, geniculados en la base; nudos y entrenudos glabros. Vainas glabras; lígulas de 0.5-3 mm, membranáceas, obtusas o truncadas, glabras; láminas de 4-27 × 0.05-0.2 cm, planas, involutas o convolutas, glabras. Panículas de 8-30 × 1-7 cm, exertas, laxas, flexuosas; ramas primarias de hasta 12 cm, adpresas a difusas; raquis glabro, escabrosa en la base de las ramificaciones; pedicelos de 0.3-1.1 cm, dispersamente escabrosos. Espiguillas con un antecio; glumas 5-nervias, lanceolado-elípticas, acuminadas o aristuladas, hialinas y verde-púrpuras, glabras; gluma inferior de 8-10.5 × 3-4 mm, gluma superior de 7.5-10 × 2.2-3.2 mm; antecio de 4.5-5.5 × 1-1.5 mm, coriáceo, obcónico, dorso giboso, ceñida debajo de la corona; lemma de 2.2-3 mm, glabra, con papilas en forma de ganchos en el tercio superior, longitudinalmente estriada; corona de 0.3-0.6 × 0.5 mm, ceñida a la base de la arista, con una hilera de macropelos de (0.2-)0.3-0.5 mm; arista de 35-45 mm, bigeniculada, escabrosa; palea de 2.2-3 mm, 2-nervia, biaquillada, surcada entre los nervios, coriácea en el ápice y dorso, navicular, puntiaguda, sobresale ligeramente de la corona, glabra; anteras de 0.5-1 mm; antopodio de 1.5-2(-2.5) mm, agudo, piloso, pelos marrón-dorados de 2-3 mm. Cariopsis de 2-2.2 × 1 mm, obovoide, glabra; embrión de 0.6 mm.
Distribución y hábitat. Nativa de Sudamérica, se encuentra en Argentina, Brasil, Chile, Uruguay y ahora en Perú (Cialdella & Arriaga 1998, Cialdella & Giussani 2002, Soreng et al. 2003). En Perú se registra en Arequipa; creciendo en campos abiertos, matorrales y asociados a los bosques de Caesalpinia spinosa (Molina) Kuntze en las formaciones de lomas de Atiquipa, de 900-1250 m de elevación.
Comentario. Piptochaetium bicolor es morfológica y filogenéticamente afín a P. brachyspermum (Speg.) Parodi, P. medium (Speg.) Torres y P. ruprechtianum É. Desv, caracterizadas por los antecios obcónicos (Cialdella & Arriaga 1998, Cialdella & Giussani 2002). P. bicolor se diferencia de P. medium y P. brachyspermum principalmente por presentar macropelos erguidos de (0.2-)0.3-0.8 mm en la corona del antecio; mientras que estructuras como los antecios de (3.5-)4.5-6.5(-7) mm y antopodios de 1-2(-2.5) mm la diferencian de P. ruprechtianum (Cialdella & Arriaga 1998). Las muestras revisadas de las formaciones de lomas presentan mínimas variaciones fuera del rango normal en comparación a Cialdella y Arriaga (1998), como: lígulas de 0.5-3mm, macropelos de la corona desde 0.2 mm y antopodio rara vez de hasta 2.5 mm. Algunos caracteres como el tamaño de los pelos de la corona podrían confundirse con los de P. brachyspermum; sin embargo, es necesario considerar una mayor cantidad de antecios y otros caracteres adicionales como el diámetro de la corona para una mejor determinación. Esta superposición de caracteres entre las dos especies ya se menciona por Parodi (1944), para lo que recomienda analizar la estructura foliar, las cuales son 6-11-fasciculada en P. bicolor y 3-fasciculada en P. brachyspermum.
Material examinado. PERÚ: Arequipa. Provincia Caravelí, Distrito Atiquipa, Lomas de Atiquipa, 15°46'12”S, 74°23'03”W, 1003 m, 12-XII- 2020, J. Muñuico et al. 718 (HSP); 15°43'50”S, 74°22'14”W, 1234 m, 13-XII-2020, J. Muñuico et al. 730 (HSP); 15°44'01”S, 74°22'22”W, 1074 m, 5-XI-2021, J. Muñuico & M. Bedoya 802 (HSP); 15°45'37”S, 74°22'33”W, 907 m, 5-XI-2021, J. Muñuico & M. Bedoya 810 (HSP).
Clave taxonómica para las especies afines a P. bicolor:
1a. Antecios de (5.5-)7.5-9.5 de largo. Antopodio de 2.5-4 mm. P. ruprechtianum É. Desv.
1b. Antecios de (3.5-)4.5-6.5(-7) de largo. Antopodio de 1-2(-2.5) mm.2
2a. Corona con macropelos erguidos de (0.2-)0.3-0.8 mm de largo, a veces con escasos ganchos y aguijones. P. bicolor (Vahl) É. Desv.
2b. Corona con ganchos y aguijones de hasta de 0.2 mm de largo (apenas sobresalen de la corona).3
3a. Antecios de (1.2-)1.5-2 mm de ancho. Corona de 0.5-0.7 mm de diámetro. Antopodio de 1.2-1.5 mm de largo. P. medium (Speg.) Torres
3b. Antecios de 1.2-1.5 mm de ancho. Corona ca. de 0.9 mm de diámetro. Antopodio de 1.9-2 mm de largo. P. brachyspermum (Speg.) Parodi
Familia: Poaceae Barnhart
Subfamilia: Panicoideae A. Braun
Supertribu: Panicodae L. Liu
Tribu: Paniceae R. Br.
Subtribu: Anthephorinae Benth.
Género: Trichachne Nees
Trichachne californica var. villosissima (Henrard) Wipff & Shaw
Figuras 1, 2E-H, 4A-G
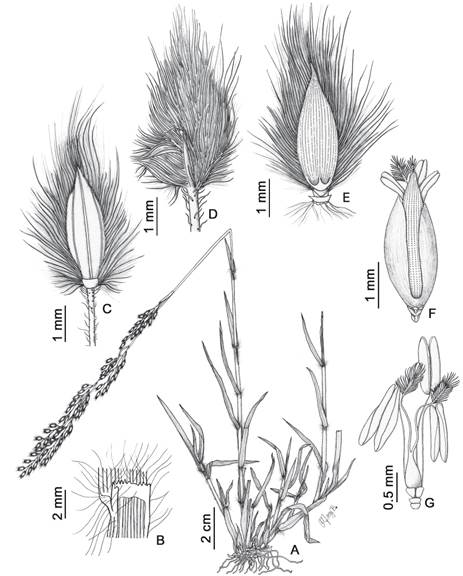
Figura 4 Trichachne californica var. villosissima. A. Hábito. B. Lígula. C. Espiguilla (vista de la gluma inferior y lemma del antecio inferior). D. Espiguilla (vista de la gluma superior). E. Antecio superior (vista dorsal). F. Antecio superior (vista ventral). G. Lodículas, androceo y gineceo. Ilustrado por Maricruz Bedoya, a partir de J. Muñuico & M. Bedoya 798 (HSP).
Phytoneuron. 2018-46:2 (2018).
=Digitaria californica var. villosissima Henrard, Monogr. Digitaria 109 (1950).
Tipo. Aruba, “Insula Aruba, Koolwijk, 1885”, W. F. R. Suringar s. n. (holotipo L-908,92-2468!; isotipo US-927379!).
Hierba perenne, de 20-60 cm de alto, brevemente rizomatosa. Culmos erguidos, generalmente ramificados y robustos en la base; nudos y entrenudos glabros. Vainas densamente pilosas; lígulas de 1.5-3 mm, membranáceas, truncadas u obtusas, erosas, glabras; láminas de 2-16 × 0.25-0.8 cm, planas, densamente pilosas (pelos de hasta 7 mm). Panículas de 9-15 × 0.5-2 cm, densas, blanquecino-violáceas; racimos de 1.5-6 cm, adpresas, ligeramente divergentes en la madurez; raquis glabro o escabroso; pedicelos de 0.5-3.5, escabrosos y glabrescentes. Espiguillas de 3-3.5 × 0.8-1.2(-1.5) mm, ovadas a ovado-lanceoladas, acuminadas, densamente pilosas, los pelos exceden a la espiguilla en 1.5-2.5 mm, con 2 antecios (inferior neutro, superior perfecto); gluma inferior de 0.3-0.5(-1) × 0.5-0.6 mm, 1-nervia, ovado-deltoidea, truncada u obtusa, hialina, glabra; gluma superior de 2.5-3.5 × 1-1.2 mm, 5-nervia, lanceolada, acuminada, densamente pilosa (pelos de 1.5-2.5(-3) mm); lemma del antecio inferior de (2.8-)3-3.5 × 1-1.2 mm, 7-nervia (dorso con 3 nervios expuestos, la central distanciado de las laterales), ovado-lanceolada, acuminada, glabra en el centro y densamente pilosa hacia los márgenes; palea del antecio inferior reducida; lemma del antecio superior de 2.5-3 × 1.3-1.5 mm, cartilaginosa, ovado-lanceolada, acuminada, rugoso-papilosa, margen hialino; palea del antecio superior de 2.2-2.5 mm, 2-nervia, ovado-lanceolada, glabra, rugoso-papilosa; lodículas 2, de 0.2 mm, truncadas; anteras de 0.6-1 mm. Cariopsis no observado.
Distribución y hábitat. Nativa de América, se encuentra en el Caribe, Venezuela, Colombia, Bolivia, Paraguay, Argentina y ahora Perú; habita en bosques secos, sabanas, quebradas, áreas disturbadas, terrenos áridos y zonas ribereñas, debajo de los 3400 m de elevación (Rúgolo de Agrasar 1974, Vega & Rúgolo de Agrasar 2002, 2005, Zuloaga et al. 2003, Lo Medico et al. 2017). En Perú crece en las formaciones de lomas de Arequipa, desde Atiquipa hasta el sur de Atico; principalmente en campos abiertos y fondo de quebradas, de 100-1200 m de elevación.
Comentarios. Trichachne californica var. villosissima ha sido tratada como Digitaria californica var. villosissima Henrard; sin embargo, Wipff y Shaw (2018) la acepta dentro del género Trichachne con respaldo en resultados tanto filogenéticos como morfológicos, las cuales son: panículas con ramas primarias dispuestas a lo largo del eje principal, espiguillas con un denso indumento de pelos cortos o largos, los pelos de la espiguilla generalmente exceden la longitud de la espiguilla, espiguilla con entrenudos de la raquila conspicuamente alargados, antecios superiores estipitados y lemma superior marrón oscuro en la madurez; mientras que Digitaria Haller se caracteriza por las panículas con ramas primarias digitadas, subdigitadas o dispuestas a lo largo del eje principal, espiguillas glabras o pubescentes, espiguillas con entrenudos de la raquilla inconspicuos, antecios superiores no estipitados y lemma superior amarillo palido, gris, púrpura, violáceo, marrón o marrón oscuro (Lo Medico et al. 2017, Wipff & Shaw 2018).
Con la inclusión de las mediciones de las muestras recolectadas en las formaciones de lomas de Perú, T. californica var. villosissima se diferenciaría de T. californica var. californica principalmente por presentar las hojas densamente pilosas (pelos de hasta 7 mm) y pelos del indumento de la espiguilla que exceden en 1.5-3 mm (nunca de 1 mm); modificándose el rango de variación de estos caracteres en comparación a Rúgolo de Agrasar (1974) y Lo Medico et al. (2017), quienes mencionan los pelos de la hoja de hasta 4 mm y los pelos del indumento de la espiguilla que exceden en 2.5-3 mm. Según Lo Medico otra diferencia entre las variedades de T. californica es la robustez de los culmos y el ancho de las hojas, sin embargo, Rúgolo de Agrasar (1974) menciona que estos caracteres son muy variables en la especie y dependen de las condiciones ambientales como humedad y nutrientes del suelo. Además, se reporta en el presente estudio las anteras de 0.6-1 mm, ligeramente mayores a los 0.5-0.6 mm del registro de Rúgolo de Agrasar (1974).
Material examinado. PERÚ: Arequipa. Provincia Caravelí, Distrito Atico, Sur de Atico - Lomas con rodal de cactus, 16°15'05”S, 73°31'04”W, 120 m, 9-XI-2021, J. Muñuico & M. Bedoya 901 (HSP); Distrito Atiquipa, Lomas de Atiquipa, 15°43'47”S, 74°22'03”W, 1184 m, 5-XI-2021, J. Muñuico & M. Bedoya 798 (HSP); Distrito Chaparra, Lomas de Capac, 15°51'25”S, 74°10'20”W, 356 m, 23-XII-2020, J. Muñuico 749 (HSP).
Clave taxonómica para los taxones afines a T. californica var. villosissima:
1a. Panícula con racimos patentes, poco espiculadas, sobre un eje de 7-10 cm, pedicelos terminales de los racimos más de 7.4 mm de largo. Espiguillas de 3-3.2 mm de largo, elipsoides o linear-lanceoladas, ápice acuminado. T. patens Swallen
1b. Panícula con racimos generalmente adpresos, densamente espiculadas, sobre un eje de 5-7 cm, pedicelos terminales de los racimos menos de 7 mm de largo. Espiguillas 3-4.5 mm de largo, ovadas, ápice abruptamente acuminado.2
2a. Vainas y láminas de las hojas glabras o escasamente pilosas. Láminas 5-13 × 0.1-0.3 cm. Indumento con pelos que exceden las espiguillas en 1 mm. T. californica (Benth.) Henrard var. californica
2b. Vainas y láminas de las hojas densamente pilosas. Láminas de (2-)6-16 × 0.25-0.8 cm. Indumento con pelos que exceden las espiguillas entre 1.5-3 mm. T. californica var. villosissima (Henrard) Wipff & Shaw
Se reporta por primera vez la presencia de Piptochaetium bicolor y Trichachne californica var. villosissima para Perú; debido a la distancia entre los registros publicados; la primera especie probablemente es introducida de los países del cono sur de Sudamérica mediante la dispersión a través de la ganadería vacuna (los cuales son abundantes en las lomas de Atiquipa), dado que según Parodi (1944), los antecios de P. bicolor presentan adaptaciones para adherirse a la piel de los animales. Por el contrario, se considera a T. californica var. villosissima como un taxón nativo, dado que presenta una amplia distribución en Sudamérica, especialmente en los países vecinos de Perú (Lo Medico et al. 2017).
Con estos nuevos registros en las formaciones de lomas, la riqueza agrostológica en éste ecosistema se incrementaría a 98 taxones a lo mencionado por Song et al. (2023). Éstos reportes son de gran importancia y relevancia para el conocimiento de las formaciones de lomas costeras; pues tal como refieren Moat et al. 2021, la fragmentación y fragilidad de este ecosistema frente a las condiciones climáticas cambiantes ponen en peligro de extinción a más de la cuarta parte de endemismos registrados y como contraste aproximadamente el 30% de la diversidad es exótica, cifra que se encuentra en incremento con el registro de P. bicolor.
Las variaciones morfológicas presentadas en el presente estudio, tanto en P. bicolor como en T. californica var. villosissima, podrían ser a causa de las condiciones ambientales peculiares de las formaciones de lomas, las que según Dillon et al. (2011) y Galán de Mera et al. (2011) dependen principalmente del suministro hídrico de las densas nieblas invernales y al ubicarse en un desierto superárido presentan limitaciones en los nutrientes del suelo. Además, algunos autores como Parodi (1944) y Rúgolo de Agrasar (1974) manifiestan otras variaciones morfológicas con muestras de otros países para los mismos taxones.
Para la flora agrostológica de Perú, el número de especies de Piptochaetium en el país se incrementaría a ocho con la incorporación de P. bicolor, (Cialdella & Arriaga 1998, Cialdella & Giussani 2002), y a 5 taxones en el caso de Trichachne con la adición de T. californica var. villosissima (Lo Medico et al. 2017).

