










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkActa Médica Peruana
On-line version ISSN 1728-5917
Acta méd. peruana vol.24 no.3 Lima Sept./dic. 2007
ARTICULOS DE REVISIÓN
Sobre expresión de genes de las enzimas de la vía glicolítica en células cancerígenas
Overexpression of genes of glycolytic pathway enzymes in cancer cells
Gustavo F. Gonzales Rengifo1, Cynthia Gonzales Castañeda2, Diego Espinosa Guerinoni2, Cristina Rojas Tubeh2.
1. Médico Endocrinólogo, Instituto de Investigaciones de la Altura y Facultad de Ciencias y Filosofía, Universidad Peruana Cayetano Heredia.
2. Facultad de Ciencias y Filosofía, Universidad Peruana Cayetano Heredia
RESUMEN
El cáncer es una importante causa de mortalidad a nivel mundial y el número de personas que se ven afectadas por esta condición va en aumento.
Se sabe que las células cancerígenas tienen una mayor actividad de la vía glicolítica respecto a las células normales, y esto se debe a una sobre expresión de los genes que codifican las enzimas que intervienen en esta ruta metabólica. Se ha visto que este metabolismo aberrante permite a las células cancerígenas cumplir su objetivo: proliferar velozmente y a su vez tener una fuente constante de energía. De esta manera se tiene una ventaja significativa con respecto a las células de los tejidos sanos. A pesar de que es ampliamente aceptada la importancia funcional de la glicólisis en el cáncer poco se conoce acerca de la influencia de la expresión genética en las elevadas tasas de esta ruta metabólica. En esta revisión se trata de recopilar los datos que sustenten el hecho de que existe una sobre expresión de los genes que codifican las enzimas de la vía glicolítica en células cancerígenas, determinar cuáles son estas enzimas y describir algunas de las técnicas empleadas en el estudio de la sobre expresión génica.
Palabras clave: glicólisis, cáncer, oncogenes, HIF-1, c-MYC
ABSTRACT
Cancer is an important cause of mortality worldwide and the number of people who are affected by it is increasing. It is known that cancer cells have greater glycolytic activity compared to normal cells. This is due to overexpression of genes which codify enzymes involved in this metabolic pathway. This aberrant metabolism allows cancer cells to proliferate quickly while using a constant source of energy. Through this, cancer cells have a significant advantage over normal tissue cells. The functional importance of glycolysis in cancer is widely accepted, but little is known about the influence of gene expression of this metabolic pathway working at high rates. In this review we compile information on the overexpression of genes codifying glycolytic pathway enzymes in cancer cells, determine which are the enzymes involved and describe some of the techniques applied in the study of gene overexpression.
Key words: glycolysis, cancer, oncogenes, HIF-1, c-MYC
INTRODUCCIÓN
Las células cancerígenas tienen una proliferación celular anormal debido a defectos en sus circuitos de regulación1. Existen más de 100 tipos distintos de cáncer, y varios subtipos de tumores que se pueden encontrar dentro de órganos específicos2. Una característica común que ha sido observada en las células cancerígenas malignas y pobremente diferenciadas es su capacidad de metabolizar glucosa a grandes velocidades3, hecho demostrado desde hace décadas basada en la observación de que los tumores presentan altas tasas de captación de glucosa y de glicólisis4. A pesar que estos cambios metabólicos no son los defectos fundamentales que causan el cáncer, ellos pueden conferir cierta ventaja en diferentes tipos de cáncer, lo que permite a la célula poder sobrevivir e invadir. Es así que los estudios recientes se ha enfocado en demostrar que varias de las alteraciones genéticas que causan el desarrollo de tumores afectan directamente a la glicólisis y a la respuesta celular a hipoxia5.
Este metabolismo aberrante que presentan las células cancerígenas sirve a la célula para poder proliferar manteniendo un suplemento constante de energía. Son varios estudios donde se observan que los tumores malignos tienen la capacidad de metabolizar la glucosa a lactato en velocidades mucho mayores que las células normales1,2,4,6,7.
La glicólisis es generalmente estudiada en un tipo de cáncer, sin embargo ya se han detectado una sobre expresión génica en 24 tipos distintos de cáncer5.
Comprender el mecanismo que desarrollan las células cancerígenas para obtener energía y así lograr su veloz proliferación; conocer nuevos rasgos celulares que caractericen a los tejidos cancerígenos y; aplicar los conceptos adquiridos en la búsqueda de nuevos blancos terapéuticos y así lograr tratamientos más efectivos son algunas de las razones que justifican el desarrollo de la siguiente revisión.
Finalmente, el objetivo que persigue este análisis es la de recopilar datos que sustenten el hecho de que existe una sobreexpresión de los genes que codifican las enzimas de la vía glicolítica en las células cancerígenas.
A su vez, esperamos poder determinar cuáles son estas enzimas de la vía glicolítica; profundizar en el estudio de la glicólisis en tejidos normales y cancerígenos y finalmente describir algunas de las técnicas que se emplean en el estudio de la sobre expresión genética.
Glicólisis y enzimas involucradas
La obtención de energía a través de la glicólisis es un mecanismo usado tanto por células normales como por células cancerígenas. El aumento de la glicólisis puede ser considerado como una adaptación o respuesta por parte de la célula cuando hay una mayor demanda de energía8. Dado que la glicólisis genera ATP, se considera como un mecanismo compensatorio cuando la fosforilación oxidativa es ineficiente, como sucede en el cáncer9.
Para entender el motivo y los mecanismos de la elevada tasa glicolítica en células cancerígenas, explicaremos primero como se realiza la glicólisis en una célula normal.
Todas las células de los mamíferos metabolizan la glucosa a piruvato por vía de la glicólisis10. Este es un sustrato único, ya que la glicólisis se puede dar tanto en ausencia de oxígeno, donde se produce lactato, como en presencia de este donde se puede metabolizar el piruvato a acetil-CoA y pasar a su oxidación completa a CO2 y agua, produciendo una gran cantidad de energía en la fosforilación oxidativa11.
En la glicólisis se degrada una molécula de glucosa (compuesta por 6 carbonos) hasta dos moléculas de piruvato (compuesta por 3 carbonos). Este proceso se da a través de 10 reacciones, catalizadas por 10 enzimas diferentes12 . Ver Figura 1.

Durante las reacciones, parte de la energía libre cedida por la glucosa es conservada en forma de ATP y NADH. De esta forma se producen 4 ATP y se gastan 2 ATP durante la glucólisis, lo que da una ganancia neta de 2 ATP12.
A continuación se muestra la vía completa de la glicólisis:
Los primeros cinco pasos de la glicólisis son denominados fase preparatoria ya que se invierten dos moléculas de ATP, elevando el contenido de energía libre de los intermediarios y convirtiendo las hexosas a un producto común que es el gliceraldehido-3-fosfato12. En esta fase participan las siguientes enzimas: hexoquinasa, fosfohexosa isomerasa, fosfofruc-toquinasa-1, aldolasa, y triosa fosfato isomerasa2.
Los últimos cinco pasos son denominados fase de beneficio donde la energía es conservada en los enlaces del ATP; el rendimiento neto es de dos moléculas de ATP. Las moléculas de ATP también permiten conservar energía12 . En esta fase participan las siguientes enzimas: gliceraldehido 3-fosfato deshidrogenasa, fosfoglicerato quinasa, fosfoglicerato mutasa, enolasa, piruvato quinasa2.
Durante la glicólisis se realizan tres tipos de transformaciones químicas: (a) Degradación del esqueleto carbonado de la glucosa (con la obtención final de piruvato), (b) Fosforilación del ADP a ATP por compuestos de fosfato de alta energía, y (c) Transferencia de átomos de un ion H+ con sus electrones hacia el NAD+, convirtiéndolo en NADH12.
Luego de que la glucosa es degradada a dos moléculas de piruvato, este puede seguir dos vías distintas en células animales13 . En condiciones anaeróbicas el piruvato se convierte a dos moléculas de lactato (fermentación láctica que se realiza en el músculo), y en condiciones aeróbicas el piruvato se convierte a dos moléculas de acetil-CoA que luego ingresa al ciclo del ácido cítrico (realizado en la mitocondria) y se degrada a CO2+H2O. Siguiendo esta vía, donde el aceptor final es el oxígeno, se obtendrán 30 ó 32 ATP, los cuales serán usados por la célula como fuente de energía para las funciones normales de la célula, como la proliferación, el crecimiento y metabolismo10 (Figuras 2 y 3).
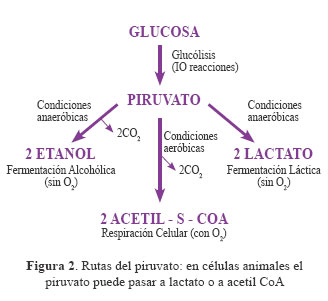
El paso de piruvato a lactato es un punto crítico en lo que se refiere al metabolismo del cáncer. La enzima que cataliza esta reacción (Piruvato ! Lactato) es la lactato deshidrogenasa (LDH). Se ha demostrado cambios en la expresión de esta enzima en células tumorales. Como consecuencia se observan cambios en la fisiología mitocondrial. Esto fue comprobado por un estudio experimental14 en el que se observó que en células tumorales deficientes de LDH hubo un aumento de la respiración mitocondrial.
Es importante mencionar que también existen otros mecanismos (aparte de la sobre expresión de enzimas) para que las células cancerígenas logren una elevada tasa glicolítica. Se ha demostrado que la expresión del transportador de glucosa 1 (GLUT-1) varía en células humanas procedentes de tejidos cancerígenos15. En el estudio citado se comprobó que aumentaba la expresión de GLUT-1 en el cáncer de mamas y que esta sobre expresión hacía que aumente la tasa de glicólisis.
Tomando en cuenta la principal característica de los tejidos cancerígenos es una elevada proliferación celular, podemos inferir que la alta tasa de glicólisis es uno de los mecanismos usados para lograrlo.
El cáncer y la glicólisis en células cancerígenas
La iniciación del cáncer se da cuando una célula no puede controlar sus procesos de división y muerte celular y por lo tanto se produce una proliferación descontrolada. Esto lleva a numerosas lesiones genéticas y epigenéticas16. Todas las células están involucradas con mecanismos de regulación del potencial de proliferación y diferenciación así como de la apoptosis1. Las dos primeras son importantes para la formación, reparación y mantenimiento de una buena función de todos los tejidos y órganos del organismo. Cuando se llega a un tamaño adecuado, las células de los tejido dejan de proliferar y diferenciarse17 .
Por lo tanto las células cancerígenas tienen dos características distintivas: (a) Se reproducen a pesar de los mecanismos de control normales, es decir, presentan un alto nivel de proliferación celular. (b) Invaden y colonizan territorios normalmente reservados para otras células. De esta forma se dan alteraciones en la morfología y fisiología, lo que produce una serie de patologías altamente dañinas para el ser humano.
Hace varias décadas, salió a luz la idea que el cáncer podría deberse a una disminución del metabolismo energético mitocondrial paralelo con un aumento en el flujo glicolítico4. Existe evidencia que sugiere que hay una relación cercana entre los cambios metabólicos y genéticos observados durante el crecimiento proliferativo6. Warburg postuló que la mayoría de tumores mostraba una alta velocidad glicolítica bajo condiciones aeróbicas2.
Las células normales utilizan el oxígeno para la producción de ATP por fosforilación oxidativa18. Cuando son privados de oxígeno, el piruvato no es metabolizado a través del ciclo de Krebs, sino que es convertido a lactato para completar los NAD y generar energía2. Durante el crecimiento tumoral, la producción de lactato a partir de glucosa se da aún en presencia de oxígeno, por lo que fue denominada como glicólisis aeróbica11, junto a un aumento de la velocidad de transporte de glucosa18.
Los tumores, a diferencia de los tejidos normales, existen en ambientes ácidos que resultan de la producción de lactato y otros ácidos2. El pH citosólico de células tumorales, sin embargo, es mantenido como en las células normales19.
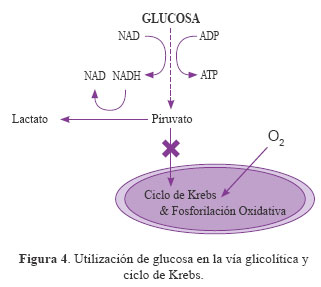
Aunque Warburg sugirió que el aumento de la glicólisis podría ser la principal causa del crecimiento tumoral, se ha visto que la eficiencia de la conversión energética mitocondrial podría ser el factor metabólico clave7, ya que el cáncer resulta de un metabolismo mitocondrial alterado11. Una gran variedad de cánceres muestran los siguientes patrones: deleción del ADN mitocondrial, contenido mitocondrial reducido, morfología mitocondrial alterada, capacidad oxidativa dañada20,21 y un aumento en la tasa glicolítica y de producción de lactato4. Este metabolismo mitocondrial alterado junto a la disminución de la actividad del ciclo de Krebs puede estar favoreciendo el crecimiento de este tipo de células7. Además, se ha observado que la frecuencia de mutaciones en el ADN mitocondrial es diez veces mayor que en el ADN nuclear22. En la Figura 4 se observa la utilización de la glucosa a través de la vía glicolítica y su producción hacia lactato en condiciones aeróbicas en células cancerígenas.
Se ha visto que el potenciamiento de la glicólisis es posible mediante diversos mecanismos como es la amplificación de genes, el aumento de su expresión, aumento de la translación, modificaciones postranslacionales y regulaciones por interacciones proteína-proteína en el citoplasma5.
De esta manera, la habilidad de mantener una velocidad incrementada de utilización de glucosa y la capacidad para sostener grandes velocidades de glicólisis bajo condiciones aeróbicas son los fenotipos bioquímicos más comunes de rápido crecimiento de células cancerígenas23. Esta elevada velocidad de catabolismo de glucosa es importante para tumores malignos, que obtienen el 50% de su energía y los precursores anabólicos para las vías biosintéticas mediante la glicólisis24.
La regulación del metabolismo glicolítico ocurre bajo una gran cantidad de vías oncogénicas y se ha visto relacionada con un aumento de agresividad del tumor16.
Esta observación sugiere que el fenotipo glicolítico juega un rol en la progresión del tumor por contribución al crecimiento o supervivencia del tumor.
A pesar que el efecto Warburg es una de las características más universales de tumores, la base molecular de este fenómeno ha sido recientemente aclarada mediante estudios25-28 que indican que el incremento de la tasa glicolítica se debería al aumento en la expresión de los genes que codifican las enzimas de la glicólisis5, provocado por la activación de factores de transcripción o oncogenes como c-MYC, USF-1, v-SRC, H-RAS o el factor inductor de hipoxia (HIF-1á)18.
La glucosa es el principal regulador de la transcripción de genes que codifican a las enzimas de la vía glicolítica, que se da mediante la estimulación de su transcripción a través del elemento de respuesta a carbohidratos (ChoRE), cuya secuencia es 5-CACGTC-3. ChoRE sirve para poder integrar señales fisiológicas mediante factores de trascripción para regular el metabolismo de glucosa18.
En la Figura 5 se observa el transporte de glucosa a través de la membrana celular por los transportadores de glucosa (GLUT-1 y GLUT-3) y el catabolismo subsiguiente de glucosa por la vía glicolítica. Se muestran los diversos puntos de regulación por el supresor de tumores p53 y von Hippel-Lindau (pVHL), así como la oncoproteína c-MYC y el factor de trascripción inductor de hipoxia 1 (HIF-1)2. Los asteriscos presentes en las diferentes enzimas indican que estas contienen elementos de respuesta a carbohidratos (ChoRE).
Las medidas de tensión de oxígeno en tumores humanos revelan una hipoxia significante2. El ambiente hostil selecciona a las células que están adaptadas a hipoxia crónica. En células normales, una respuesta crítica a hipoxia es la inducción a factor de trascripción inductor de hipoxia, HIF-1, que promueve la trascripción de genes asociados al metabolismo29. Este se une a la secuencia de ADN e incrementa la expresión de genes que codifica a las enzimas glicolíticas y de esta manera promueve la adaptación celular para reducir la disponibilidad de oxígeno mediante el aumento de la ingesta de glucosa y el aumento de la glicólisis30 .
Lo que produce la hipoxia en células cancerígenas es inducir fisiológicamente la expresión de genes de la vía glicolítica mediante los sitios de unión a HIF-118. De esta manera, HIF-1 se une a los genes cuyos sitios de unión tengan la secuencia central 5-RCGTG-325 para de esta manera poder promover la adaptación celular necesaria para poder disminuir la disponibilidad de oxígeno mediante un aumento de la glucosa y glicólisis29. Esto se va a lograr mediante un aumento de la expresión de genes que codifican las enzimas glicolíticas como la aldolasa A, la enolasa 1, la lactato deshidrogenada A, la fosfofructoquinasa L, fosfoglicerato quinasa 1, y piruvato quinasa M, así como el gen del factor de crecimiento vascular endotelial (VEGF)2 y el transportador de glucosa, GLUT-131. Sin embargo, no solo es necesario del sitio de unión sino que también se necesita que HIF-1 interactúe con otros factores de trascripción unidos a sitios adyacentes25-28. Como resultado de las alteraciones genéticas y la hipoxia intratumoral, el HIF-1 es sobre-expresado en la mayoría de los cánceres humanos comunes25-28.

Las células proliferativas expresan al VEGF que va a estimular la angiogénesis para dar una mayor perfusión y mantener de esa manera la oxigenación28, la cual es regulada negativamente por c-MYC18.
Además de las alteraciones en la tensión de oxígeno, los cambios en las concentraciones de glucosa también activan varios genes de las enzimas glicolíticas a través del elemento de respuesta a carbohidratos, el cual encaja a los sitios de unión en la secuencia para MYC y HIF-132, 5-RCGTG-3, que sirven para activar la trascripción18. La activación de las vías de HIF-1 pueden mediar las respuestas adaptativas a hipoxia e hipoglicemia en células cancerígenas. Alternativamente, la activación de oncogenes o la pérdida de los supresores de tumores por mutaciones somáticas, pueden llevar directamente a alteraciones no fisiológicas del metabolismo celular y proveer una ventaja selectiva en ambientes metabólicos hostiles2.
El oncogene c-MYC se encuentra activado en una variedad de vías importantes para el control del crecimiento celular y tumorigénesis18 y codifica a un factor de trascripción que lo que hace es hetero-dimerizar a otro oncogene (MAX) para que se pueda unir a la secuencia 5-CACGTG-32. De esta manera c-MYC estimula la captura de glucosa y también al metabolismo así como va a activar la maquinaria del ciclo celular necesaria para la proliferación celular18. La expresión de c-MYC deja a las células susceptibles a la muerte por varios estímulos33 y es un blanco directo para mutaciones oncogénicas, mientras que la expresión del factor inductor de hipoxia 1 (HIF-1) es regulada indirectamente vía la ganancia de funciones de mutaciones en oncogenes y la pérdida de funciones de mutaciones en genes supresores de tumores26,27.
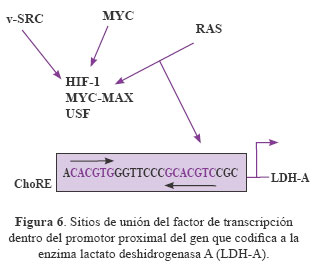
En la Figura 6 se puede observar los sitios de unión del factor de trascripción dentro del promotor proximal del gen que codifica la enzima lactato deshidrogenasa A (LDH-A). La caja E (5-CACGTG-3) es el centro de consenso del elemento de respuesta a carbohidratos (ChoRE) y se une con los sitios de unión para la trascripción del HIF-1, MYC-MAX y USF. MYC y HIF-1 pueden unirse a los elementos cis directamente, mientras que v-SRC y H-RAS activado potencian la actividad de HIF-1 y otros factores que se unen a estos elementos y activan la glicólisis.
Para el crecimiento de tumores en condiciones hipóxicas, se necesita de un alto flujo glucolítico, y según lo observado, muchas de las células transformadas muestran esta alta tasa de glicólisis29. Este flujo es controlado principalmente por la 6-fosfofructo-2-quinasa, siendo la fructosa-2,6-bifosfato (Fru-2,6-P2) el activador alostérico más poderoso34, y de esta manera se obtiene un control de la vía glicolítica. La 6-fosfofructo-2-quinasa/fructosa-2,6-bifosfatasa es una enzima bifuncional cuya función es catalizar la síntesis y degradación de Fru-2,6-P2 y de esa manera regular el metabolismo de carbohidratos35.
Se han visto cuatro genes implicados en la codificación de las diferentes isoformas de la familia de la 6-fosfofructo-2-quinasa, la PKFB 1-429. Estas isoformas muestran diferencias en la distribución de sus tejidos y además tienen propiedades cinéticas en respuesta a las señales efectores alostéricas, hormonales y de factor de crecimiento36. El gen PKFB 3 se encuentra ubicado en diversos lugares y es expresada continuamente en tejidos proliferativos37,38, en líneas celulares transformadas39, y en varios tumores40. El producto de este gen tiene el cociente de la actividad quinasa/fosfatasa bien alta41 con lo que indica que en los tejidos donde va a ser expresada, se mantendrán los niveles de fructosa-2,6-bifosfato y las altas tasas de glicólisis serán sostenidas29.
La principal consecuencia de una elevada tasa de glicólisis en células tumorales es que los carbonos de glucosa son convertidos principalmente a lactato3 a altas velocidades mayores que en células normales2, por lo tanto dejan de ser la principal fuente de carbonos para la respiración aeróbica. En las células tumorales que son capaces de consumir cantidades limitadas de oxígeno, la glutamina es el principal sustrato oxidable que entra a un ciclo de Krebs truncado2. El aumento en esta velocidad en el transporte de glucosa, depende de los niveles elevados de la enzima hexoquinasa42. Esta enzima participa en el primer paso de la vía glicolítica donde convierte la glucosa a glucosa 6-fosfato, el cual es el intermediario fosforilado inicial de la vía glicolítica2.
La glucólisis en algunos tipos de células de mamíferos se ha reportado que lo controlan la hexokinasa (HK) y la fosfofructokinasa-I (PFK-I) entre un 70 % y un 30%, respectivamente43. La enzima hexokinasa ha atraído la atención debido a que está involucrada en la iniciación y mantenimiento de altas tasas de catabolismo de glucosa en tumores con rápido crecimiento44, y porque se ha visto que el gen que codifica esta enzima (del tipo II), se encuentra amplificada en líneas celulares de hepatoma y como consecuencia se encuentra marcadamente elevada en los tumores45. El rol de la hexokinasa ha sido tema de gran investigación para poder entender la base molecular de su fenotipo glicolítico aberrante, y ha sido considerado como un blanco potencial para el arresto del crecimiento celular de tumores. Se ha logrado observar que en comparación con células normales, la actividad de la hexokinasa es marcadamente elevada en la glicólisis en tumores en avanzado crecimiento24, y que en células deficientes del HIF-1 á, la expresión de la hexoquinasa II se encuentra marcadamente disminuida46.
En las células tumorales AS-30D (de rata) se determina que este porcentaje cambia, de tal forma que la HK junto con el transportador de glucosa controlan 71%, 4 % la PFK-I y 25% el bloque de enzimas conformado por la aldolasa, TIM, GAPDH, PGK, PGM, ENO, PYK, LDH y la demanda de ATP43. La redistribución del control en las células tumorales AS-30D se origina aparentemente debido al aumento en la actividad de todas las enzimas de la glucólisis (con respecto a hepatocito), pero específicamente de la hexokinasa y la fosfofructokinasa-I45, que son las enzimas que incrementan en mayor proporción su actividad (124 y 22 veces respectivamente), lo que ocasiona un aumento en las concentraciones de sus productos, la glucosa 6-fosfato y la fructosa 1,6-bifosfato en 5 y 250 veces respectivamente, trayendo como consecuencia un incremento en la velocidad de glicólisis de 20 veces43.
Los altos niveles de glicólisis no solo proveen de ATP para la gran demanda bioenergética de las células tumorales, sino que también provee de precursores y reduce a los equivalentes para la síntesis de macromoléculas19. En varios tejidos, la síntesis de ácidos grasos ocurre a bajas velocidades, ya que los lípidos son adquiridos vía la circulación para proveer las necesidades de las células vegetativas no proliferativas. En contraste, la nueva síntesis de ácidos grasos ocurre a grandes velocidades en tejidos tumorales47. En la mayoría de las células tumorales, casi todos los ácidos grasos son derivados de la síntesis de novo a pesar de tener un suplemento abundante de lípidos extracelulares. La gran tasa de síntesis de ácidos grasos en células altamente proliferativas, provee de energía a la biogénesis de la membrana. Es posible que el incremento del metabolismo de la glucosa contribuya a la proliferación de células tumorales por promover a la síntesis de ácidos grasos48.
En células que han pasado a una conversión glicolítica (han pasado de un modo de metabolismo oxidativo a uno glicolítico), gran cantidad de la demanda bioenergética es suministrada a través de la producción glicolítica de ATP, y el piruvato entra a un ciclo truncado donde el citrato es exportado preferentemente al citosol vía el transportador tricarboxilato49. Una vez en el citosol, el citrato es cortado por la ATP citrato liasa para producir acetil Co-A citosólico y regenerar el oxalacetato. El acetil-CoA es el requisito para la síntesis endógena de ácidos grasos, colesterol e isoprenoides, así como las reacciones de acetilación que modifica a las proteínas19.
Para completar este ciclo, el oxalacetato es reducido a malato, el que puede regresar a la mitocondria, reciclando carbono y reduciendo equivalentes en la mitocondria. La conversión de oxalacetato a malato citosólico es dirigido por el alto radio de NADH/NAD+ citosólico presente en las células glicolíticas50. El malato puede entrar en la matriz mitocondrial y ahí ser convertido a oxalacetato para completar el ciclo. La conversión de NAD+ a NADH provee un mecanismo continuo para preservar el potencial de la membrana mitocondrial y sostener un gran radio NADH/NAD+ mitocondrial que mantenga el ciclo truncado en estado reprimido. Además, la actividad enzimática del ATP-citrato liasa está equilibrada para afectar tanto a la lipogénesis dependiente de glucosa y a la bioenergética celular19.
La ATP-citratoliasa (ACL) es una enzima homotetramérica con una expresión en gran cantidad de tejidos, lo que exhibe una regulación transcripcional coordinada con otras enzimas en la vía lipogénica48. Los niveles de ACL aumentan en respuesta a señales que comunican un estado nutricional, activa la liberación y metabolismo de glucosa celular, y estimula un crecimiento anabólico19.
Las vías de señalización que contribuyen al fenotipo glicolítico y juegan un papel importante en la tumorigénesis, pueden también llevar a incrementar los niveles y/o actividad de la ACL48. Estas vías pueden explicar en parte el hecho que la actividad de la ACL se ha visto significantemente elevada en carcinomas de mamas y vejiga versus tejidos normales de mamas y vejiga51.
La conversión de glicólisis aeróbica parece ser capaz de promover el crecimiento tumoral mediante la estimulación de síntesis de lípidos y mediante la supresión de la diferenciación de la célula tumoral. Estas propiedades pueden contribuir a la selección de una conversión a glicólisis anaeróbica durante la progresión de tumores in vivo (el efecto Warburg) y al parecer los inhibidores de ACL pueden tener un rol potencial en suprimir el crecimiento de tumores malignos19.
En los últimos años se ha visto que solo se ha estudiado la glicólisis en unos cuantos cánceres52, y por lo general se ha visto el estudio de la inducción de la 6-fosfofructo-2-quinasa en líneas celulares de varios cánceres humanos53. En la Tabla 1 se puede observar los diversos genes y su expresión en 24 tipos de cáncer. Se ha observado que en cánceres de cerebro, nódulo linfático y próstata existe una sobre expresión de genes de las enzimas de la vía glicolítica. Sin embargo, los cánceres de colon y glándula mamaria parecen tener otro tipo de mecanismos ya que no aparentan tener una máxima sobre expresión39. Aún así, la glicólisis se sigue manteniendo afectada.
Todas las sobre expresiones de genes se observan que ocurren en la glicólisis. No se ha encontrado enninguna otra vía bioquímica un patrón consistente de la sobre expresión génica, ni siquiera en el ciclo del ácido cítrico la cual también provee de energía a los tejidos cancerígenos5.
Técnicas para evaluar sobre expresión de genes
a. Análisis de Northern Blot.
El método de Northern Blot continúa siendo, a pesar del enorme desarrollo de nuevas tecnologías como el PCR en Tiempo Real, la herramienta estándar para el análisis cualitativo y cuantitativo de niveles de ARN mensajero (ARNm) en una determinada célula o tejido de interés.

Esta técnica consiste básicamente en la extracción de ARN total (ARN del tipo mensajero, ribosomal y de transferencia). Posteriormente se separa por tamaño en un gel de agarosa el tipo de ARN que se desee estudiar y se transfiere por adhesión a una membrana de nylon. Finalmente se procede a hibridizar este ARN con una determinada sonda o secuencia de ADN o ARN que contenga a su vez un reportero (un marcador radioactivo por ejemplo) que permita identificar, visualizar e identificar la hibridización de ambos fragmentos de ácidos nucleicos y así determinar la posición y la cantidad de la región de interés54.
Las principales desventajas que tiene esta metodología implican:
- Fragilidad del ARN, dado que es muy susceptible a degradarse si no se toman las debidas precauciones tales como el uso de material libre de ARNasas y el almacenamiento a temperaturas adecuadas (-70ºC).
- No tiene la sensibilidad con la que cuentan otras técnicas más recientes.
- El desarrollo de sondas de hibridización para más de una región es un proceso complicado.
Para el análisis específico de tejidos cancerígenos la metodología implica la separación del tejido sano, la posterior extracción de ARN total y su posterior análisis como se ha descrito anteriormente. El objetivo central en este caso es detectar y cuantificar la cantidad de ARN mensajero de interés, es decir, aquel que contenga la secuencia específica que codifique la enzima que se desea estudiar. Por último se procede a comparar el patrón obtenido con el tejido sano44.
b. Transcripción reversa y PCR (RT-PCR).
Este método combina la reacción en cadena de la polimerasa (PCR) con el proceso de transcripción reversa, es decir, la conversión del ARN a ADN. El PCR es una técnica desarrollada durante los años ochenta por Kary Mullis y se basa en el descubrimiento de la actividad biológica a altas Tº de las ADN polimerasas encontradas en bacterias termófilas56.
Tiene como propósito generar un gran número de copias de un fragmento de ADN de interés in vitro. La concentración de copias del gen aumenta exponencialmente, ya que en cada ciclo se copian las dos cadenas del ADN. Si se inicia la reacción con una copia del gen y éste es copiado en el primer ciclo, se ha de tener dos copias. En el segundo ciclo se a de concluir con la formación de cuatro copias, etc.
La transcripción reversa, tanto utilizando ARN total como ARNm, es la etapa clave en el proceso de RT-PCR57. Existen varios factores que influencian la eficiencia del proceso, tales como la acción de la enzima que va a llevar a cabo esta reacción y la presencia de estructuras secundarias en el ARN58.
Las reverso transcriptasas virales, tales como M-MLV y AMV, han sido las enzimas de elección durante muchos años59. Estas enzimas, sin embargo, son termolábiles, y no pueden llevar a cabo la transcripción reversa cuando existen estructuras secundarias en el ARN. En este caso, se suelen utilizar transcriptasas reversas termoestables, que pueden llevar a cabo la reacción a 55-70 ºC, permitiendo la desnaturalización de las estructuras secundarias, y aumentando la eficiencia global de la reacción60.
El método de RT-PCR permite medir la expresión génica mediante una técnica alternativa al Northern Blot. Es más sensible y requiere menos ARN. Sin embargo, la presencia de ARN o ADN contaminante, tanto en las muestras, como en el laboratorio y reactivos, debe ser evitada con el fin de impedir la aparición de productos inespecíficos61.
En el caso del estudio de tejidos cancerígenos se procede realizando la extracción de ARN del tejido, se efectúa la transcripción reversa y finalmente se lleva a cabo el PCR. Los resultados son finalmente comparados con los tejidos sanos62.
c. PCR en Tiempo Real.
Esta técnica se basa en la detección de un reportero fluorescente en cada ciclo de la reacción, por lo que el monitoreo es en tiempo real.
La señal de fluorescencia se incrementa de una forma directamente proporcional a la cantidad de producto de amplificación. Esto nos permite realizar análisis basándonos en la amplificación durante los primeros ciclos de la reacción y no al final de la misma, como con el PCR convencional.
Los métodos de detección utilizando la técnica de PCR convencional presentan varios inconvenientes, siendo el más serio el riesgo de resultados falsos positivos por contaminación con amplicones63. Como alternativa, se desarrolló la técnica del PCR en Tiempo Real, la cual es más rápida, sensible, precisa, práctica y, en principio, libre de problemas de contaminación64.
A grandes rasgos el procedimiento para el análisis de tejidos cancerígenos consiste en comparar los niveles de expresión de la secuencia de interés, expresados en número de copias, entre el tejido cancerígeno y el sano65.
d. Secuenciamiento.
El secuenciamiento es un procedimiento que requiere de la acción de la ADN polimerasa que sintetiza la cadena de ADN en presencia de nucleótidos trifosfato (dNTPs) y de análogos de los nucleótidos trifosfato (ddNTPs) acoplados a un compuesto radioactivo o cromogénico. Los ddNTPs interrumpen el proceso de síntesis del ADN ya que no permiten la elongación de la cadena sintetizada. La reacción de polimerización en estas condiciones genera fragmentos de ADN de tamaño variable, los cuales son separados por electroforesis y visualizados 66.
Tiene como principal ventaja ser la herramienta por excelencia de análisis molecular ya que permite determinar con exactitud la secuencia de nucleótidos en una región específica o de interés y así detectar la presencia de mutaciones, re-arreglos genéticos y zonas polimórficas. Por otro lado, la gran desventaja que tiene esta técnica es el elevado costo de los equipos necesarios para llevarla a cabo67.
Para el estudio de tejidos cancerígenos el secuenciamiento ha sido una herramienta muy útil dado que ha permitido, por ejemplo, detectar un patrón de metilación del ADN característico de las células cancerígenas hepáticas con respecto a las células sanas del mismo órgano45.
CONCLUSIONES
Se ha observado que en las células cancerígenas existe un cambio del metabolismo aeróbico69. A pesar de la aceptación de la importancia de la glicólisis en el cáncer se conoce poco acerca de cómo esta influencia en la expresión de genes de las enzimas sobre expresadas en este metabolismo5. La actividad incrementada y la expresión de estas enzimas son identificadas en tumores ya que más del 50% de su energía y sus precursores anabólicos derivan de la vía glicolítica47.
Se ha considerado como blanco para tratamiento al HIF-126,27, a la HK-II29, y a la ATP citrato liasa19. Aun así, todos estos actúan conjuntamente con otras proteínas y no son las únicas maneras de crear una sobre-expresión de genes. Es por eso que se dificulta en la manera de encontrar un tratamiento eficaz para detener el crecimiento proliferativo y el desarrollo de las células cancerígenas.
Las técnicas para la sobre expresión de genes son diversas. Cada una de las presentadas en esta revisión cuenta con una serie de ventajas y desventajas lo cual le delega la responsabilidad al investigador de elegir la que más se adecue a sus objetivos experimentales y posibilidades económicas.
REFERENCIAS BIBLIOGRÁFICAS
1. Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100:57-70. [ Links ]
2. Dang CV, Semenza GL. Oncogenic alterations of metabolism. Trends Biochem Sci. 1999; 24:68-72. [ Links ]
3. Mathupala S. P., Rempel A, Pedersen P. Glucose catabolism in cancer cells. The Biol Chem. 2001; 276:43407–43412. [ Links ]
4. Warburg O. On the origin of cancer cells. Science. 1956; 123:309–314. [ Links ]
5. Altenberga B, Greulichb K.O. Genes of glycolysis are ubiquitously overexpressed in 24 cancer classes. Genomics. 2004; 84:1014– 1020 [ Links ]
6. Ramanathan A, Wang C, Schreiber SL. Perturbational profiling of a cell-line model of tumorigenesis by using metabolic measurements. Proc Natl Acad Sci U S A. 2005; 102:5992-7 [ Links ]
7. Schulz T, Thierbach R, Voig A, Drewes G, Mietzner B, Steinberg P, Pfeiffer A, Ristow M. Induction of oxidative metabolism by mitochondrial frataxin Inhibits cancer growth. The J Biol Chem 2006; 281:977–981. [ Links ]
8. Vogt AM, Elsasser A, Nef H, Bode C, Kubler W, Schaper J. Increased glycolysis as protective adaptation of energy depleted, degenerating human hibernating myocardium. Mol Cell Biochem. 2003; 242:101-7. [ Links ]
9. Carew JS., Huang P. Mitochondrial defects in cancer. Mol Cancer 2002; 1:9 [ Links ]
10. Costello L, Franklin R. Why do tumour cells glycolyse?: From glycolysis through citrate to lipogenesis. Mol Cell Biochem. 2005; 280:1–8. [ Links ]
11. Gatenby RA, Gillies RJ. Why do cancers have high aerobic glycolysis? Nat Rev Cancer. 2004; 4:891-9. [ Links ]
12. Nelson D, Cox M. Lehningers principles of biochemistry. Fourth Edition. WH Freeman and Company. New York.2005. 521-525.
13. Bui T, Thompson C. Cancers sweet tooth. Cancer Cell 2006; 9:419-420. [ Links ]
14. Fantin VR, St-Pierre J, Leder P. Attenuation of LDH-A expression uncovers a link between glycolysis, mitochondrial physiology, and tumor maintenance. Cancer Cell. 2006; 9:425-34. [ Links ]
15. Rivenzon-Segal D, Boldin-Adamsky S, Seger D, Seger R, Degani H. Glycolysis and glucose transporter 1 as markers of response to hormonal therapy in breast cancer. Int J Cancer 2003; 107:177-82. [ Links ]
16. Elstrom, R.L., Bauer, D.E., Buzzai, M., Karnauskas, R., Harris, M.H., Plas, D.R., Zhuang, H., Cinalli, R.M., Alavi, A., Rudin, C.M., and Thompson, C.B. Akt stimulates aerobic glycolysis in cancer cells. Cancer Res 2004; 64:3892–3899. [ Links ]
17. Pardo G, Hernández P, Delgado R. La apoptosis y la senescencia celular: mecanismos supresores de tumores. Rev Cub Med 2005. 44:1-12 [ Links ]
18. Osthus R.C., Shim H., Kim S., Li Q., Reddy R., Mukherjee M., Xu Y., Wonsey D., Lee L.A., Dang, C.J. Deregulation of glucose transporter 1 and glycolytic gene expression by c-Myc. Biol. Chem. 2000; 275:21797-21800. [ Links ]
19. Hatzivassiliou G, Zhao F, Bauer DE, Andreadis C, Shaw AN, Dhanak D, Hingorani SR, Tuveson DA, Thompson CB. ATP citrate lyase inhibition can suppress tumor cell growth. Cancer Cell. 2005; 8:311-321. [ Links ]
20. Cuezva J.M., Krajewska M., de Heredia ML., Krajewski S., Santamaria G., Kim H. The bioenergetic signature of cancer: a marker of tumor progression. Cancer Res. 2002; 62:6674-6681. [ Links ]
21. Rossignol R., Gilkerson R., Aggeler R., Yamagata K., Remington S.J., Capaldi R.A. Energy substrate modulates mitochondrial structure and oxidative capacity in cancer cells. Cancer Res. 2004; 64:985–993 [ Links ]
22. Verma M, Kagan J, Sidransky D, Srivastava S. Proteomic analysis of cancer-cell mitochondria. Nat Rev Cancer 2003; 3:789-795. [ Links ]
23. Eigenbrodt, E., Fister, P., Reinacher, M. New perspectives in carbohydrate metabolism in tumor cells. In: Reitner, R. (ed). Regulation of carbohydrate metabolism. CRC Press. 1985; 2:141-179.
24. Mathupala S, Rempel A, Pedersen P. Glucose catabolism in cancer cells. Isolation, sequence, and activity of the promoter for type II hexokinase. The J Biol Chem 1995; 270:16918-16925. [ Links ]
25. Semenza G.L. Hypoxia-inducible factor 1: master regulator of O2 homeostasis. Curr. Opin. Genet. Dev. 1998; 8:588-594. [ Links ]
26. Semenza GL, Artemov D, Bedi A, Bhujwalla Z, Chiles K, Feldser D, Laughner E, Ravi R, Simons J, Taghavi P, Zhong H. The metabolism of tumours: 70 years later. Novartis Found Symp. 2001; 240:251-60. [ Links ]
27. Semenza GL. Hypoxia-inducible factor 1: oxygen homeostasis and disease pathophysiology. Trends Mol Med. 2001a; 7:345-50. [ Links ]
28. Semenza GL. Targeting HIF-1 for cancer therapy. Nat Rev Cancer 2003; 3:721-32. [ Links ]
29. Obach M, Navarro-Sabate A, Caro J, Kong X, Duran J, Gómez M, Perales JC, Ventura F, Rosa JL, Bartrons R. 6-Phosphofructo-2-kinase (PFKFB3) gene promoter contains hypoxia- inducible Factor-1 binding sites necessary for transactivation in response to hypoxia. The J Biol chem. 2004; 279:53562–53570. [ Links ]
30. Minchenko A., Leshchinsky I., Opentanova I., Sang N., Srinivas V., Armstead V., Caro J.J. Hypoxia-inducible factor-1-mediated expression of the 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase-3 (PFKFB3) gene.Its possible role in the Warburg effect. Biol. Chem. 2002; 277, 6183–6187 [ Links ]
31. Chen C., Pore N., Behrooz A., Ismail-Beigi F., Maity AJ. Regulation of glut1 mRNA by hypoxia-inducible factor-1. Interaction between H-ras and hypoxia. Biol. Chem. 2001; 276:9519–9525 [ Links ]
32. Grandori C., Eisenman R. N. Myc target genes. Trends Biochem. Sci. 1997; 22:177–181 [ Links ]
33. Shim H, Chun YS, Lewis BC, Dang CV. A unique glucose-dependent apoptotic pathway induced by c-Myc. Proc Natl Acad Sci U S A. 1998; 95:1511-1516. [ Links ]
34. Pilkis S.J., Claus T.H., Kurland I.J., Lange A.J. 6-Phosphofructo-2-kinase/fructose-2,6-bisphosphatase: a metabolic signaling enzyme. Annu. Rev. Biochem. 1995; 64, 799–835 [ Links ]
35. Hue L., Rousseau G.G. Fructose 2,6-bisphosphate and the control of glycolysis by growth factors, tumor promoters and oncogenes. Adv Enzyme Regul. 1993; 33, 97–110 [ Links ]
36. Okar DA, Manzano A, Navarro-Sabate A, Riera L, Bartrons R, Lange AJ. PFK-2/FBPase-2: maker and breaker of the essential biofactor fructose-2,6-bisphosphate. Trends Biochem Sci. 2001; 26:30-35. [ Links ]
37. Hamilton JA, Callaghan MJ, Sutherland RL, Watts CK. Identification of PRG1, a novel progestin-responsive gene with sequence homology to 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase. Mol Endocrinol. 1997;11:490-502. [ Links ]
38. Riera L., Manzano A., Navarro-Sabate A., Perales J.C., Bartrons R. Insulin induces PFKFB3 gene expression in HT29 human colon adenocarcinoma cells. Biochim. Biophys. Acta. 2002; 1589:89–92 [ Links ]
39. Chesney J. An inducible gene product for 6-phosphofructo-2-kinase with an AU-rich instability element: role in tumor cell glycolysis and the Warburg effect, Proc Natl Acad Sci 1999; 3047– 3052. [ Links ]
40. Atsumi, T., Chesney, J., Metz, C., Leng, L., Donnelly, S., Makita, Z., Mitchell, R., Bucala, R. Cancer Res 2002; 62:5881–5887 [ Links ]
41. Sakakibara R, kato M, Okamura N, Nakagawa T, Komada Y, Tominaga N, Shimojo M, Fukasawa M. Characterization of a human placental fructose-6-phosphate, 2-kinase/fructose-2,6-bisphosphatase. J Biochem (Tokyo)1997; 122:122-128. [ Links ]
42. Rempel A., Mathupala S.P., Griffin C.A. Hawkins A.L., Pedersen P. Glucose catabolism in cancer cells: amplification of the gene encoding type II hexokinase. Cancer Res 1996; 56:2468–2471. [ Links ]
43. Hernandez M, Sanchez R. Control de la glucólisis en células tumorales de rápido crecimiento Memorias del XIV Congreso de Bioenergética y Biomembranas. 2005.
44. Arora K, Pedersen P. Functional significance of mitochondrial bound hexokinase in tumor cell metabolism. Evidence for preferential phosphorylation of glucose by intramitochondrially generated ATP. J Biol Chem. 1988; 263:17422-17428. [ Links ]
45. Goel A, Mathupala S, Pedersen P. Glucose Metabolism in Cancer. J Biol Chem. 2003; 278:15333-15340. [ Links ]
46. Iyer N.V., Kotch LE, Agani F, Leung SW, Laughner E, Wenger RH, Gassmann M, Gearhart JD, Lawler AM, Yu AY, Semenza GL. Cellular and developmental control of O2 homeostasis by hypoxia-inducible factor 1 alpha. Genes Dev. 1998; 12:149–162 [ Links ]
47. Kuhajda, F.P. Fatty-acid synthase and human cancer: new perspectives on its role in tumor biology. Nutrition. 2000; 16:202–208. [ Links ]
48. Towle, H.C., Kaytor, E.N., and Shih, H.M. Regulation of the expression of lipogenic enzyme genes by carbohydrate. Annu. Rev. Nutr. 1997; 17:405–433. [ Links ]
49. Baggetto, L.G. Deviant energetic metabolism of glycolytic cancer cells. Biochimie. 1992; 74, 959–974. [ Links ]
50. Bauer, D.E., Harris, M.H., Plas, D.R., Lum, J.J., Hammerman, P.S., Rathmell, J.C., Riley, J.L., and Thompson, C.B. Cytokine stimulation of aerobic glycolysis in hematopoietic cells exceeds proliferative demand. FASEB J. 2004; 18: 1303–1305. [ Links ]
51. Turyn, J., Schlichtholz, B., Dettlaff-Pokora, A., Presler, M., Goyke, E., Matuszewski, M., Kmiec, Z., Krajka, K., and Swierczynski, J. Increased activity of glycerol 3-phosphate dehydrogenase and other lipogenic enzymes in human bladder cancer. Horm. Metab. Res. 2003; 35:565–569. [ Links ]
52. Zu XL, Guppy M. Cancer metabolism: facts, fantasy, and fiction. Biochem Biophys Res Commun 2004; 313:459–465. [ Links ]
53. Haberkorn U. FDG uptake, tumor proliferation and expresión of glycolysis-associated genes in animal tumor models. Nucl. Med. Biol. 1994; 21:827– 834. [ Links ]
54. Porchet N, Aubert JP. Northern blot analysis of large mRNAs. Methods Mol Biol. 2000; 125:305-12. [ Links ]
55. Coutinho, Cláudia Malheiros, Bassini, Alessandra Simões, Gutiérrez, Leonardo Guilhermino et al. Genetic alterations in Ki-ras and Ha-ras genes in juvenile nasopharyngeal angiofibromas and head and neck cancer. Sao Paulo Med. J. 1999;117:113-120. [ Links ]
56. Mullis KB, Faloona FA. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods Enzymol. 1987; 155:335-50 [ Links ]
57. Bustin SA. Quantification of mRNA using real-time reverse transcription PCR (RT-PCR): trends and problems. J of Mol Endocrinol. 2002; 29:23-39[ [ Links ]STANDARDIZEDENDPARAG]
58. Phillips JK, Lipski J. Single-cell RT-PCR as a tool to study gene expression in central and peripheral autonomic neurones. Auton Neurosci 2000; 86:1-12 [ Links ]
59. Menendez-Arias L. Molecular basis of fidelity of DNA synthesis and nucleotide specificity of retroviral reverse transcriptases. Prog Nucleic Acid Res Mol Biol 2002; 71:91-147. [ Links ]
60. Zhang YJ, Pan HY, Gao SJ. Reverse transcription slippage over the mRNA secondary structure of the LIP1 gene. Biotechniques 2001; 31:1286-1290. [ Links ]
61. Roth CM. Quantifying gene expression. Curr Issues Mol Biol 2002; 4:93-100. [ Links ]
62. Rubie C, Kempf K, Hans J, Su T, Tilton B, Georg T, Brittner B, Ludwig B, Schilling M. Housekeeping gene variability in normal and cancerous colorectal, pancreatic, esophageal, gastric and hepatic tissues. Mol Cell Probes 2005; 19:101-109 [ Links ]
63. Peirson SN, Butler JN, Foster RG. Experimental validation of novel and conventional approaches to quantitative real-time PCR data analysis. Nucleic Acids Res 2003;31:e73. [ Links ]
64. Kubista M, Andrade JM, Bengtsson M, Forootan A, Jonak J, Lind K, Sindelka R, Sjoback R, Sjogreen B, Strombom L, Stahlberg A, Zoric N. The real-time polymerase chain reaction. Mol Aspects Med 2006; 27:95-125. [ Links ]
65. Radonic A, Thulke S, Mackay IM, Landt O, Siegert W, Nitsche A. Guideline to reference gene selection for quantitative real-time PCR. Biochem Biophys Res Commun 2004; 313:856–62. [ Links ]
66. Huang GM. High-throughput DNA sequencing: a genomic data manufacturing process. DNA Seq 1999; 10:149-53. [ Links ]
67. Franca LT, Carrilho E, Kist TB. A review of DNA sequencing techniques. Q Rev Biophys 2002; 35:169-200. [ Links ]
68. Espinoza JR. La hélice dorada y la medicina molecular. Revi Diagnóstico 2000; 39: 294-301. [ Links ]
69. Keleg S, Buchler P, Ludwig R, Buchler MW, Friess H. Invasion and metastasis in pancreatic cancer. Mol Cancer 2003;2:14. [ Links ] Review.
CORRESPONDENCIA
Cynthia Gonzales-Castañeda
e-mail: dah_182@yahoo.com
