










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa Médica Peruana
versión On-line ISSN 1728-5917
Acta méd. Peru vol.36 no.2 Lima abr./jun. 2019
ARTÍCULO ORIGINAL
Efecto antigenotóxico de las semillas de Salvia hispanica "chia" utilizando la prueba de Allium cepa
Antigenotoxic effect of Salvia hispanica "chia" seeds using the Allium cepa test
Daniel López-Rojas1,a, Lizzie Becerra-Gutiérrez1,2,b, Cristian Díaz-Vélez1,3,c
1 Facultad de Medicina, Universidad de San Martin de Porres. Chiclayo, Perú.
2 Laboratorio de Investigación de Inmunología y Virología, Hospital Regional Lambayeque. Chiclayo, Perú.
3 Hospital Nacional Almanzor Aguinaga Asenjo. Chiclayo, Perú.
a Médico cirujano, b Doctora en Microbiología, c Médico con maestría en Epidemiología Clínica
RESUMEN
Objetivo: El objetivo fue evaluar el efecto antigenotóxico de las semillas de Salvia hispanica expuestas al efecto genotóxico de la azida de sodio utilizando la prueba de Allium cepa. Materiales y métodos: Estudio experimental, las unidades de análisis fueron células meristemáticas de 40 bulbos de Allium cepa. Se dispuso 5 grupos de trabajo: control negativo, control positivo, pretratamiento, simultáneo y postratamiento, además de dos concentraciones (10% y 20%). Las raíces se expusieron a los tratamientos por un total de 48 horas. Resultados: Se observó una disminución significativa del índice mitótico (IM) del control negativo (23,7) respecto al control positivo (3,4) confirmando la acción genotóxica de la azida de sodio. En el grupo pretratamiento, la reducción de daño (RD%) fue de -151,2% y -59,2% señalando su nulo papel desmutagénico. En el grupo postratamiento y simultáneo, la RD% fue de 37,6%, 67,2% y 59,2%, 64,8% respectivamente para ambas concentraciones indicando su actividad bioantimutagénica dependiente de la concentración. Sin embargo, a las concentraciones evaluadas no se halló significancia estadística. Conclusión: Las semillas de Salvia hispanica expuestas al efecto genotóxico de la azida de sodio utilizando la prueba de Allium cepa no tienen un efecto antigenotóxico a las concentraciones de 10% y 20%.
Palabras clave: Antimutagénicos; Aberraciones cromosómicas; Índice mitótico; Salvia; Daño del ADN (fuente: DeCS-BIREME).
ABSTRACT
Objective: The objective was to assess the antigenotoxic effect of Salvia hispanica seeds exposed to the genotoxic effect of sodium azide using the Allium cepa test. Materials and Methods: This is an experimental study. The analyzed units were meristem cells from forty Allium cepa bulbs Five working groups were available: negative controls, positive controls, pre-treatment, simultaneous, and post-treatment, and two concentrations (10% and 20%) were used. Bulbs were exposed to treatment for a total 48-hour time. Results: A significant reduction in the mitotic index (MI) of negative controls (23.7) was observed, compared to positive controls (3.4), confirming the genotoxic action of sodium azide. In the pre-treatment group, damage reduction values (DR%) were -151.2% and -59.2%, indicating their absent demutagenic role. In both posttreatment and simultaneous groups, DR% values were 37.6 %, 67.2 %, 59.2 %, and 64.8 % for both concentrations, respectively, indicating their concentration-dependent bioantimutagenic activity. However, no statistically significant differences were found between the concentrations tested. Conclusion: Salvia hispanica seeds exposed to the genotoxic effect of sodium azide using the Allium cepa test have no antigenotoxic effect at 10% and 20% concentrations.
Keywords: Antimutagenic agents; Chromosome aberrations; Mitotic index; Salvia; DNA damage (source: MeSH NLM).
INTRODUCCIÓN
El cáncer es una de las principales causas de morbilidad y mortalidad a nivel mundial; la Organización Mundial de la Salud estima que los casos anuales aumentarán de 14 millones en 2012 a 22 millones dentro de los próximos 20 años [1]. Según la Organización Panamericana de la Salud, en la región de las Américas el número de muertes aumentará de 1,2 millones en 2008 hasta 2,1 millones en 2030 [2,3]. En Perú, se diagnostican más de 45 000 casos nuevos de cáncer cada año [4]. Mientras que, en Lambayeque se ha observado un aumento de la prevalencia de los casos de algunos tipos de cáncer, prioritarios en salud pública, entre los años 2007 al 2010; representando estos casos, la pérdida de 4 780,4 años de vida saludables perdidos (AVISA), en el 2013 [3,5].
El incremento en la aparición del cáncer puede estar relacionado por el aumento de la frecuencia de alteraciones al ADN, producto de la exposición del ser humano a agentes genotóxicos, capaces de ocasionar alteraciones en el material genético originando efectos adversos en el organismo [6,7]. Es importante señalar que un exceso de radicales libres, aunque sean productos normales del metabolismo celular, puede tener efectos genotóxicos; por lo que toma relevancia el estudio del valor de los suplementos dietarios con antioxidantes, definidos como un grupo de enzimas, minerales y vitaminas que ayudan a eliminar los radicales libres impidiendo la oxidación de diversas sustancias cuyas reacciones pueden provocar alteraciones fisiológicas causantes de diversas enfermedades [8-10].
Por otro lado, el uso de plantas medicinales por sus propiedades curativas y/o preventivas, una práctica habitual en aproximadamente el 79% de la población mundial, brinda la posibilidad de encontrar compuestos químicos con actividad biológica [9]. Dentro de estos compuestos, se considera que las plantas medicinales son una de las mayores fuentes de antioxidantes naturales. Así mismo, se estima que el 60% de drogas anticancerígenas provienen directa o indirectamente de las plantas medicinales [7]. Salvia hispanica L., es una planta herbácea de la familia de las lamiáceas, localmente conocida por sus usos medicinales, y con el nombre común de "chía", proveniente de los pueblos indígenas de las épocas precolombinas y aztecas. Sus semillas son ricas en ácidos grasos omega 3, 6 y 9, fibras y una gran cantidad de antioxidantes, por lo que recientemente se ha ampliado su uso para varios beneficios para la salud; por ejemplo, se han encontrado estudios previos donde se demuestra la actividad antitumoral de extractos de esta especie vegetal en adenocarcinoma de mama murina y como modificador de la resistencia a vinblastina en células cancerígenas de mama in vitro [11-13].
Sin embargo, en la búsqueda bibliográfica no se ha encontrado la descripción de un posible efecto antigenotóxico, por lo que en este estudio se planteó como objetivo general evaluar tal efecto utilizando la prueba de Allium cepa, cuyo modelo permite observar el daño en el ADN en células eucariotas, por lo que los resultados pueden ser extrapolables para todas las plantas y animales [14].
MATERIALES Y MÉTODOS
Tipo y diseño de estudio
Estudio de tipo experimental, prospectivo, con grupo control, medición basal y aleatorización.
Material vegetal
Se usaron semillas orgánicas de Salvia hispanica "chía", adquiridas de la empresa Sierra & Selva® de la ciudad de Lima, y molidas mediante una licuadora. Los principales compuestos antioxidantes bioactivos presentes en este licuado fueron quercetina, kaempferol, myricetina, ácido cafeico y ácido clorogénico [11,15]. Se sumergieron 25 g de semillas molidas de Salvia hispanica en 250 ml de agua destilada y se dejaron reposar por 24 horas, obteniéndose de esta manera una concentración de 0,1 g/ml equivalente al 10%; así mismo, se sumergieron 50 g de semillas molidas de Salvia hispanica en 250 ml de agua destilada y se dejaron reposar por 24 horas, obteniéndose de esta manera una concentración de 0,2 g/ml, equivalente al 20%.
Para inducir daño al ADN, se empleó como agente mutágeno al compuesto azida de sodio (NaN3) a una concentración de 0,05%, que fue donado de la Universidad Agraria de la Selva de la ciudad de Huánuco.
Medición del efecto genotoxico con Allium cepa
Se usaron bulbos de Allium cepa como unidades experimentales y se realizaron 5 repeticiones por cada tratamiento y por cada concentración, sumando un total de 40 unidades experimentales utilizadas. Se incluyeron bulbos procedentes de un solo tipo de cosecha, y se excluyeron bulbos con características muy diferentes a sus pares en lo que respecta a tamaño y color.
A temperatura ambiente (19 a 23°C), se colocó cada unidad experimental en un vaso descartable con agua potable de modo que sólo su disco radicular quedó en contacto con el líquido y se esperó hasta que las raíces tengan un tamaño de 2-3 cm para luego exponerlas a las sustancias a estudiar.
Posteriormente, se distribuyó las unidades experimentales en los siguientes grupos de estudio:
• Control negativo: se expusieron las raíces de cada bulbo en un nuevo vaso descartable con agua potable durante 48 horas.
• Control positivo: se expusieron las raíces de cada bulbo en un nuevo vaso descartable con azida de sodio durante 48 horas.
• Grupo pretratamiento: se expusieron las raíces de cada bulbo en un nuevo vaso descartable con Salvia hispanica durante 24 horas, luego de las cuales se trasladó el bulbo a un nuevo vaso descartable con azida de sodio de modo que sus raíces quedaron expuestas a la sustancia durante 24 horas más.
• Grupo simultáneo: se expusieron las raíces de cada bulbo en un nuevo vaso descartable con solución de Salvia hispanica y azida de sodio al mismo tiempo durante 48 horas.
• Grupo postratamiento: se expusieron las raíces de cada bulbo en un nuevo vaso descartable con azida de sodio durante 24 horas, luego de las cuales se trasladó el bulbo a un nuevo vaso descartable con Salvia hispanica de modo que sus raíces quedaron expuestas a la solución durante 24 horas más.
Las células de Allium cepa fueron extraídas de la región meristemática de las raíces de cada bulbo después de 48 horas, con modificaciones en la técnica descrita por Akinboro et al. [16]:
Primero, una raíz de cada bulbo fue cortada con ayuda de una hoja de bisturí con una longitud de 1 cm desde su ápice, para ser sumergida en un tubo de ensayo en solución de fijación (1:3 ácido acético:etanol) por 30 minutos. La raíz fijada fue enjuagada en agua destilada por 15 minutos. Luego, fue sometida a hidrólisis ácida para extraer el material genético (1N HCl a 60°C por 12 minutos). La raíz hidrolizada fue enjuagada en agua destilada por 15 minutos. Después, los cromosomas fueron teñidos con orceína acética 1,5% durante 25 minutos.
Después de ello, la raíz fue nuevamente cortada con una longitud de 5 mm desde su ápice para extraer la región meristemática. Finalmente, se colocó una gota de gelatina fenicada sobre el material vegetal cubriéndolo con una laminilla cubreobjetos, inmediatamente después se aplicó la técnica squash de aplastamiento con la ayuda de papel toalla para lograr una mejor distribución de las células y se procedió a observar al microscopio a aumentos de 40x y 60x. Para observar células al microscopio con aumento de 100x se necesitó sellar los bordes de las laminillas cubreobjetos con resina transparente y cubrirlas con una gota de aceite de inmersión.
Fueron analizadas las células de la región meristemática de las raíces de un total de 40 bulbos de Allium cepa mediante un microscopio óptico. Durante el análisis, las células fueron divididas en interfase, profase, metafase, anafase y telofase; además se cuantificó el número de aberraciones cromosómicas (AC).
Recolección de datos
Los datos obtenidos fueron recogidos con ayuda de una ficha de recolección de datos que consistió en dos secciones, una de células normales donde se recogió información del número de células en cada fase de la mitosis (interfase, profase, metafase, anafase, telofase), índice mitótico y porcentaje de reducción del índice mitótico, y otra sección de aberraciones cromosómicas, donde se contó el número de células aberrantes por cada tipo de aberración cromosómica (micronúcleos, puentes cromosómicos, células binucleadas, células apoptóticas, células anucleadas, núcleo a la periferia, cromosomas adherentes, c-mitosis) y el porcentaje de reducción de daño.
Para obtener el índice mitótico (IM), el número de células en división (profase, metafase, anafase, telofase) fue dividido por el número total de células analizadas [17].

El porcentaje de reducción del índice mitótico (RIM%) fue calculado usando la siguiente fórmula [17]:

El porcentaje de reducción de daño (RD%) fue obtenido usando la frecuencia de aberraciones cromosómicas (AC) en el siguiente cálculo [17]:
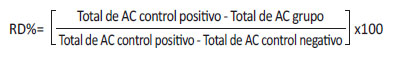
Análisis estadístico
Los datos recogidos fueron procesados para su análisis y ajuste estadístico y asimismo para su respectiva interpretación, para lo cual se utilizó el software IBM SPSS Statistics for Windows, Version 23.0. ® (Armonk, NY: IBM Corp). Se realizó estadística descriptiva para media e intervalo de confianza por cada grupo. Usando la prueba de Kolmogorov-Smirnov para normalidad, resultó que la base de datos no tenía una distribución normal por lo que se usó la prueba de Kruskal-Wallis con el cual se confirmó que los datos provenían de poblaciones diferentes. Por último, se utilizó la prueba de Games-Howel para comparación de pares de medias. Se consideró un nivel de significancia p<0,05 y un nivel de confianza de 95%.
Aspectos éticos
Se contó con el permiso institucional de la Universidad San Martin de Porres – Filial Norte – Chiclayo. Se utilizaron datos obtenidos del procesamiento de material biológico vegetal, por lo que no se requirió del consentimiento informado de sujeto alguno.
Se usaron medidas y material de bioseguridad y almacenamiento aislado para el manejo de los materiales en este estudio, especialmente del agente genotóxico (azida de sodio). Se procedió a la esterilización de todo material de laboratorio que estuvo en contacto con el agente genotóxico (azida de sodio) para evitar daño a terceras personas y al medio ambiente. Estas consideraciones aseguraron la calidad de la información obtenida en este estudio.
RESULTADOS
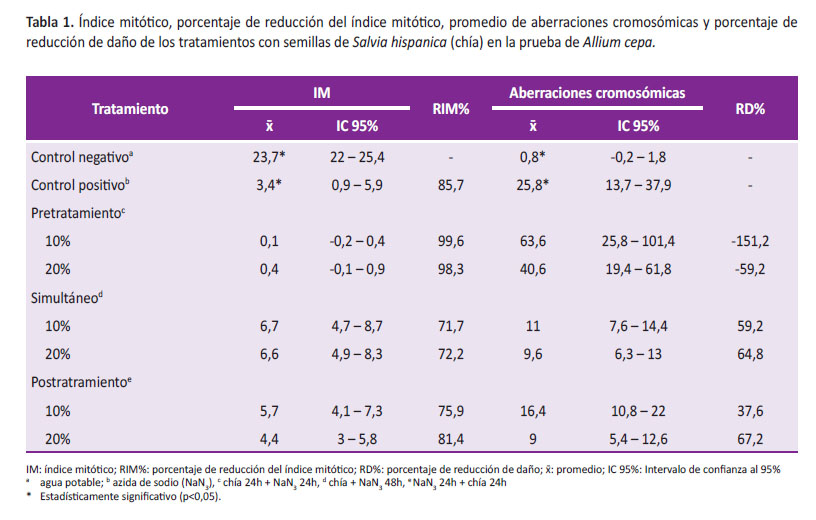
En el grupo control positivo se observó un índice mitótico (IM) por debajo del control negativo, con una reducción del 85,7% (p<0,05) (Tabla 1). Por otro lado, la frecuencia de aberraciones cromosómicas del control positivo fue mayor que la encontrada en el control negativo (p<0,05) (Figura 1).
En forma similar, se evidencio que en ambas concentraciones del grupo pretratamiento, los índices mitóticos estuvieron por debajo del control positivo; sin embargo, cuando la concentración aumentaba se observó una leve mejoría en el IM, disminuyendo el porcentaje de reducción del IM, aunque ésta no fue estadísticamente significativa (p>0,05). La frecuencia de aberraciones cromosómicas a ambas concentraciones fue mayor que la encontrada en el control positivo evidenciándose una nula reducción de daño (p>0,05). En forma similar a lo observado en el IM, al aumentar la concentración se evidenció una disminución de la frecuencia de aberraciones cromosómicas y una mejora en el porcentaje de reducción de daño, que no fue estadísticamente significativa (p>0,05).
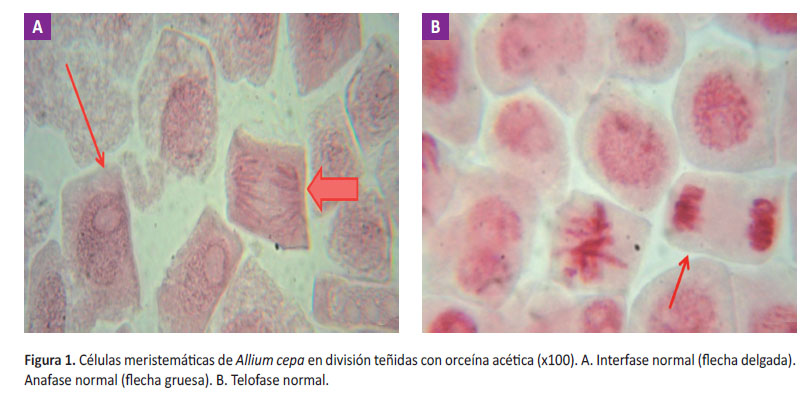
Cuando se realizó la comparación entre el grupo simultáneo y postratamiento, se notó que a ambas concentraciones hubo una mejoría en el IM respecto al control positivo evidenciándose un menor porcentaje de reducción de IM respecto al control positivo, no estadísticamente significativo (p>0,05). Al aumentar la concentración se evidenció una leve disminución de su IM como un leve aumento de su porcentaje de reducción del IM. Por otro lado, la frecuencia de aberraciones cromosómicas a ambas concentraciones fue menor que el control positivo, aumentando el porcentaje de reducción de daño, sin embargo, estas diferencias no fueron significativas (p>0,05). Además, se observó que al aumentar la concentración la frecuencia de aberraciones cromosómicas disminuía y el porcentaje de reducción de daño aumentaba (p>0,05) (Figura 2).
DISCUSIÓN
En la presente investigación no se pudo demostrar, estadísticamente, un efecto protector de las semillas de Salvia hispanica (chía) frente al daño al material genético (aberraciones cromosómicas), usando la prueba de Allium cepa. Sin embargo, las aberraciones cromosómicas fueron menores a una concentración más alta de Salvia hispanica. Esto indicaría un potencial beneficio de sus componentes antioxidantes, como quercetina, kaempferol, myricetina, ácido cafeico y ácido clorogénico, para disminuir las aberraciones cromosómicas. Por lo tanto, a concentraciones más altas de Salvia hispanica que las del presente estudio, se podría obtener resultados estadísticamente significativos.
Se decidió utilizar las concentraciones de 25 g y 50 g de semillas de Salvia hispanica tomando como ejemplo al ensayo clínico controlado de Nieman et al. donde a estas concentraciones se quiso evaluar la efectividad de las semillas de Salvia hispanica para promover la pérdida de peso y disminuir los factores de riesgo de enfermedad de adultos con sobrepeso. Además, estas concentraciones se encuentran en el rango que muchos otros ensayos clínicos controlados usaron para evaluar diferentes beneficios de las semillas de Salvia hispanica [18-24].
La prueba de Allium cepa ha sido considerada por muchos autores como de bajo costo y de fácil manejo, una de sus ventajas es la presencia de un sistema enzimático oxidasa, que es esencial para las evaluaciones de mutagenicidad. Por lo tanto, mientras que otras pruebas, por ejemplo, la prueba de Ames, requiere la adición de la mezcla S9 (fracción S9 del hígado de rata), la prueba de Allium cepa presenta capacidad metabólica para activar promutágenos en mutágenos sin la adición de un sistema metabólico exógeno para evaluar esta clase de contaminantes. Sin embargo, cuando el sistema enzimático oxidasa de las plantas superiores se compara con las enzimas del citocromo P-450 de mamíferos, el complejo enzimático de las plantas presenta una concentración baja y una limitación en la especificación del sustrato [25]. Sin embargo, los resultados de los bioensayos de plantas superiores no deben descartarse, ya que un producto químico capaz de inducir daños cromosómicos en las plantas también puede ofrecer riesgos a otros grupos de organismos vivos, ya que el material dañado es el ADN, que es común a todos los organismos [26].
Según los resultados observados, en todos los grupos de trabajo donde se usó azida de sodio se evidenció una disminución del índice mitótico y un aumento de aberraciones cromosómicas confirmando el poder genotóxico de esta sustancia al producir una inhibición de la replicación normal y ocasionando daños a nivel molecular, resultados similares se encontraron en el estudio de Ragunathan et al. donde se reporta que el efecto genotóxico de la azida de sodio se puede deber a la formación de un gas tóxico de hidrógeno de azida con la generación de iones de azida cuando ésta se disuelve en agua o a la acumulación de peróxido que causa toxicidad [14]. El ADN es particularmente sensible al daño ocasionado por las especies reactivas de oxígeno (como el peróxido) causando deterioro de las membranas lipídicas, proteínas y ácidos nucleicos [27].
En el presente estudio se evaluó el modo de acción de las semillas de Salvia hispanica en la protección al material genético frente a un daño conocido. Según estudios, existen dos mecanismos de acción por medio de los cuales se puede evaluar esta citoprotección: la desmutagénesis, referida a la capacidad para bloquear el daño al ADN, principalmente mediante la absorción del agente genotóxico actuando preferentemente en el medio extracelular; y la bioantimutagénesis, referida a la capacidad para mejorar los sistemas de reparación de división celular, actuando por lo tanto, intracelularmente [6].
En la investigación realizada, en el grupo pretratamiento, se evaluó la actividad desmutagénica, encontrándose que hubo menor replicación celular (menor índice mitótico) y mayor frecuencia de aberraciones cromosómicas que las ocasionadas por la azida de sodio, sin embargo, entre las concentraciones estudiadas, se observó una mejoría en el índice mitótico y una disminución de frecuencia de aberraciones cromosómicas cuando la concentración aumentaba. Estos resultados contrastan con los hallazgos de Fedel-Miyasato et al. donde el extracto metanólico de Shinus terebinthifolius demostró tener una actividad desmutagénica presumiblemente por la inactivación directa de los componentes del genotóxico a nivel del citoplasma [7].
Los resultados del presente estudio también difieren a lo encontrado por Glińska et al., donde se evidenció un poder desmutagénico de Brassica oleracea var. capitata f. rubra debido a la acción quelante de sus compuestos fenólicos [27]. Asimismo, los resultados negativos contrastan con el estudio de Rosas-Ramirez et al. donde se evaluaron los polisacáridos del mucílago de las semillas de Salvia hispanica para descubrir compuestos moduladores de la resistencia a quimioterapéuticos en células de cáncer de mama, obteniéndose una actividad moderada en potenciar en alrededor de cinco veces la respuesta al fármaco vinblastina en líneas celulares con multidrogo resistencia por sobre expresión de la P-glicoproteina. Por lo que, además de los antioxidantes, los polisacáridos de Salvia hispanica podrían tener un efecto significativo desde una perspectiva terapéutica [13]. Por lo tanto, la pobre actividad desmutagénica de las semillas de Salvia hispanica frente al daño producido por la azida de sodio probablemente se deba a la poca concentración evaluada.
En el grupo postratamiento y simultáneo se evaluó actividad bioantimutagénica, encontrándose que hubo un aumento en el índice mitótico y una disminución de la frecuencia de aberraciones cromosómicas comparadas con las ocasionadas por la azida de sodio. Resultados similares también se observaron en el estudio de Fedel-Miyasato et al. donde el extracto metanólico de Schinus terebinthifolius también demostró tener actividad bioantimutagénica reparando el daño a través del ciclo celular, debido posiblemente a su gran cantidad de fitoquímicos como fenoles y flavonoides [7]. Los resultados de este estudio también coincidieron con los de Mauro et al. donde se evidenció que la inulina podía modular las enzimas de las células vegetales para mejorar la reparación del daño genético producido por una exposición previa a un genotóxico [17]. Otro estudio con resultados similares a los del presente trabajo fue el de Sultan et al. donde se observó que el efecto protector pudo ser debido a las propiedades antioxidantes de los flavonoides del extracto de Capparis spinosa [28]. Una investigación de Akinboro et al. demostró que el extracto acuoso de Myristica fragrans tenía un potencial antimutagénico gracias a la actividad antioxidante atribuida a sus compuestos fenólicos y polifenólicos [29]. En un estudio de Sondhi et al. se encontró que los brasinoesteroides de las hojas de Aegle marmelos disminuían la frecuencia de aberraciones cromosómicas posiblemente debido a la regulación de la expresión de genes, ácidos nucleicos y proteínas [30]. Finalmente, se encontró resultados coherentes con el estudio de Carnier et al. donde al evaluar la dieta suplementada con harina de Salvia hispanica sobre el desarrollo de la caquexia del cáncer y la respuesta inflamatoria de los tejidos, no previno los efectos de los tumores en modelos animales [31]. Esto se puede deber a la pobre actividad desmutagénica hallada también en el presente estudio. Por lo tanto, la buena actividad bioantimutagénica de las semillas de Salvia hispanica se debe probablemente a los antioxidantes que contiene.
Las semillas de Salvia hispanica tienen una gran cantidad de compuestos polifenólicos, desde 0,511 a 0,881 mg/g dependiendo del método de extracción [32], con capacidad antioxidante como el ácido cafeico (0,003-0,156 mg/g), el ácido clorogénico (0,046-0,102 mg/g) y los flavonoides, entre los que se encuentran la quercetina (0,150-0,268 mg/g), la myrcetina y el kaemferol [11,15,32]. Estos antioxidantes actúan a través de varios mecanismos incluyendo la neutralización de radicales libres, la modulación de expresión de genes, la inhibición de enzimas oxidativas y la estimulación de enzimas antioxidantes [7,27]. La quercetina es un poderoso antioxidante capaz de prevenir la oxidación de grasas, proteínas y ADN, y estas propiedades antioxidantes son significativamente más efectivas que otros flavonoides [15]. Por otro lado, existen estudios donde se ha demostrado que a dosis altas algunos flavonoides como la quercetina pueden actuar como prooxidantes y exhibiendo efectos genotóxicos en modelos bacterianos [33,34]. La actividad prooxidante de la quercetina es probablemente atribuible a su capacidad para autooxidarse o convertirse en metiluros intermedios como orto-semiquinona y orto-quinona/quinona a través de la oxidación enzimática, lo que puede contribuir a la generación de especies reactivas de oxígeno y glutatión [35].
Sin embargo, se deben considerar varios puntos para comprender por qué la quercetina es mutagénica in vitro, pero no mutagénica/carcinogénica in vivo. En primer lugar, el metabolismo de la quercetina in vivo, particularmente el efecto de primer paso, reduce significativamente el potencial de efectos adversos mediados por la quercetina después del consumo oral. Por ejemplo, la O-metilación da como resultado la eliminación de las características estructurales esenciales que subyacen a la mutagenicidad relacionada con la quercetina. En segundo lugar, existen varios mecanismos de protección in vivo para limitar la actividad pro-oxidante de las pequeñas cantidades de quercetina no metabolizada disponible sistémicamente, y combinados con la baja biodisponibilidad de la aglicona de quercetina actúan al unísono para limitar la posibilidad de que ocurra cualquiera de los efectos adversos que se observan in vitro. Además, dada la muy poca biodisponibilidad de aglicona de quercetina, no se espera que estos mecanismos de protección estén saturados incluso a niveles más altos de exposición a la quercetina. Los niveles de dosis en los estudios a largo plazo en animales en los que no se informaron efectos adversos toxicológicamente significativos apoyan la adición de quercetina a la alimentación a dosis estimadas de exposición que se aproximan a las ingestas de quercetina natural de la dieta por parte de los consumidores con una alta ingesta de frutas y verduras (es decir, 200–500 mg/día) [35].
Además, las semillas de Salvia hispanica contienen pequeñas cantidades de vitamina E, que es ampliamente aceptada como uno de los antioxidantes más potentes, sobretodo su isoforma tocotrienol, su propiedad antioxidante se atribuye al grupo hidroxilo del anillo aromático de tococromanoles, que dona hidrogeno para neutralizar los radicales libres o especies reactivas de oxígeno [15,36]. En los últimos años se ha indicado fuertemente que los tocotrienoles pueden prevenir/inhibir eficazmente el crecimiento de diferentes tipos de cáncer (cerebro, mama, cuello uterino, hígado, pulmón, próstata, piel, páncreas, estómago, etc) debido a su capacidad para modular varias dianas moleculares involucradas en la proliferación, supervivencia, invasión, angiogénesis y metástasis de células cancerosas [37].
Por consiguiente, la leve actividad desmutagénica y la buena actividad bioantimutagénica de las semillas de Salvia hispanica evidenciadas en este estudio se deban probablemente a la gran cantidad de antioxidantes que tienen. A pesar de evidenciar la actividad bioantimutagénica de las semillas de Salvia hispanica, con las concentraciones evaluadas no se encontró diferencia estadística.
Como limitación se tuvo que las raíces de los bulbos de Allium cepa podrían no crecer en el tiempo establecido. Fue viable ya que se contó con la sustancia genotóxica (azida de sodio), las unidades experimentales (bulbos de Allium cepa) y con las semillas de Salvia hispanica, con el ambiente de laboratorio adecuado para el desarrollo del estudio, con el tiempo libre necesario y con el presupuesto suficiente para la realización de la presente investigación.
Se puede concluir que las semillas de Salvia hispanica expuestas al efecto antigenotóxico de la azida de sodio utilizando la prueba de Allium cepa no tienen un efecto antigenotóxico a las concentraciones de 10% y 20%, la azida de sodio produce un efecto genotóxico evidenciado por la disminución del índice mitótico y el aumento de las aberraciones cromosómicas y que, entre todos los grupos estudiados, el tratamiento simultáneo tiene un mejor porcentaje de reducción de índice mitótico y porcentaje de reducción de daño dosis dependiente.
Para futuros trabajos de investigación se recomienda aumentar el número de repeticiones por cada grupo de trabajo. También se sugiere aumentar el número de unidades experimentales por cada repetición en cada grupo de trabajo. Asimismo, se aconseja aumentar el tiempo de exposición a las sustancias a evaluar, así como, su medición de pH. Además, se recomienda trabajar con concentraciones más altas que las utilizadas en el presente estudio.
Contribución de autoría: DLR y LBG han participado en la concepción y diseño del artículo, recolección de datos, análisis e interpretación de datos. Además, DLR realizó la redacción del artículo y aportó el material de estudio, mientras que CDV hizo la revisión crítica del artículo, la aprobación de la versión final y la asesoría estadística.
Fuentes de financiamiento: Autofinanciado.
Conflictos de interés: Los autores declaran no tener algún conflicto de interés.
REFERENCIAS BIBLIOGRÁFICAS
1. Organización Mundial de la Salud. Cáncer [Internet]. Geneva: OMS; 2015 [citado 22 de marzo de 2015]. Disponible en: https://www.who.int/es/news-room/fact-sheets/detail/cancer [ Links ]
2. Ramos WC, Venegas DR, Medina JL, Guerrero PC, Cruz A. Análisis de la situación del cáncer en el Perú 2013 [Internet]. Lima: Ministerio de Salud del Perú; 2013 [citado 30 de marzo de 2015]. Disponible en: https://www.dge.gob.pe/portal/docs/asis_cancer.pdf [ Links ]
3. Díaz Vélez C. Carga de Enfermedad de los Cánceres Prioritarios en la Red Asistencial de Lambayeque, Perú 2013. Rev Venez Oncol. 2016;28(4):204-15. [ Links ]
4. Ministerio de Salud del Perú. Esperanza de Vencer el Cancer [Internet]. Lima: Ministerio de Salud del Perú; 2013 [citado 23 de marzo de 2015]. Disponible en: https://www.minsa.gob.pe/Especial/2013/cancer/index.asp [ Links ]
5. Díaz Vélez C, Peña Sánchez R. Factores pronóstico de sobrevida de los cánceres prioritarios en el seguro social ESSALUD Lambayeque Perú. Rev Venez Oncol. 2012;24(3):202-16. [ Links ]
6. Abrevaya X. ¿Qué es la genotoxicidad? [Internet]. Buenos Aires, IntraMed; 2008 [citado 1 de mayo de 2015]. Disponible en: https://www.intramed.net/contenidover.asp?contenidoID=47111 [ Links ]
7. Fedel-Miyasato LE, Formagio AS, Auharek SA, Kassuya CA, Navarro SD, Cunha-Laura AL, et al. Antigenotoxic and antimutagenic effects of Schinus terebinthifolius Raddi in Allium cepa and Swiss mice: a comparative study. Genet Mol Res GMR. 2014;13(2):3411-25. [ Links ]
8. Zamora JD. Antioxidantes: Micronutrientes en Lucha por la Salud. Rev Chil Nutr. 2007;34(1):17-26. [ Links ]
9. Martínez García N. Evaluación del efecto quimioprotector de la decocción de la raíz de Jatropha dioica en ratones albinos suizos cepa ICR mediante Ensayo cometa in vivo. [Tesis de Maestría]. San Agustín Tlaxiaca, Hidalgo, México: Universidad Autónoma del Estado de Hidalgo; 2013. [ Links ]
10. Civetta MTM de, Civetta JD. Carcinogénesis. Salud Pública México. 2011;53(5):405-14. [ Links ]
11. Mohd Ali N, Yeap SK, Ho WY, Beh BK, Tan SW, Tan SG. The Promising Future of Chia, Salvia hispanica L. J Biomed Biotechnol. 2012;2012:171956. [ Links ]
12. Espada CE, Berra MA, Martinez MJ, Eynard AR, Pasqualini ME. Effect of Chia oil (Salvia Hispanica) rich in omega-3 fatty acids on the eicosanoid release, apoptosis and T-lymphocyte tumor infiltration in a murine mammary gland adenocarcinoma. Prostaglandins Leukot Essent Fatty Acids. 2007;77(1):21-8. [ Links ]
13. Rosas-Ramírez DG, Fragoso-Serrano M, Escandón-Rivera S, Vargas-Ramírez AL, Reyes-Grajeda JP, Soriano-García M. Resistance-modifying Activity in Vinblastine-resistant Human Breast Cancer Cells by Oligosaccharides Obtained from Mucilage of Chia Seeds (Salvia hispanica). Phytother Res PTR. 2017;31(6):906-14. [ Links ]
14. Tedesco SB, Laughinghouse HD IV. Bioindicator of Genotoxicity: The Allium cepa Test. En: Srivastava J. Environmental Contamination. 1ra edición. Londres: IntechOpen Limited; 2012. p. 137-56. [ Links ]
15. Hernández LM. Mucílago de Chía (Salvia hispanica): microestructura, caracterización físico-química y aplicaciones en la industria alimentaria [Tesis Doctoral]. Santiago de Compostela, España: Universidad de Santiago de Compostela; 2012. [ Links ]
16. Akinboro A, Mohamed KB, Asmawi MZ, Othman AS, Ying TH, Maidin SM. Mutagenic and antimutagenic assessment of methanol leaf extract of Myristica fragrans (Houtt.) using in vitro and in vivo genetic assays. Drug Chem Toxicol. 2012;35(4):412-22. [ Links ]
17. Mauro MO, Pesarini JR, Marin-Morales MA, Monreal MTFD, Monreal ACD, Mantovani MS, et al. Evaluation of the antimutagenic activity and mode of action of the fructooligosaccharide inulin in the meristematic cells of Allium cepa culture. Genet Mol Res GMR. 2014;13(3):4808-19. [ Links ]
18. Nieman DC, Cayea EJ, Austin MD, Henson DA, McAnulty SR, Jin F. Chia seed does not promote weight loss or alter disease risk factors in overweight adults. Nutr Res. 2009;29(6):414-8. [ Links ]
19. Ho H, Lee AS, Jovanovski E, Jenkins AL, Desouza R, Vuksan V. Effect of whole and ground Salba seeds (Salvia Hispanica L.) on postprandial glycemia in healthy volunteers: a randomized controlled, dose-response trial. Eur J Clin Nutr. 2013;67(7):786-8. [ Links ]
20. Tavares Toscano L, Tavares Toscano L, Leite Tavares R, da Oliveira Silva CS, Silva AS. Chia induces clinically discrete weight loss and improves lipid profile only in altered previous values. Nutr Hosp. 2014;31(3):1176-82. [ Links ]
21. Toscano LT, da Silva CSO, Toscano LT, de Almeida AEM, Santos A da C, Silva AS. Chia flour supplementation reduces blood pressure in hypertensive subjects. Plant Foods Hum Nutr. 2014;69(4):392-8. [ Links ]
22. Vuksan V, Jenkins AL, Dias AG, Lee AS, Jovanovski E, Rogovik AL, et al. Reduction in postprandial glucose excursion and prolongation of satiety: possible explanation of the long-term effects of whole grain Salba (Salvia Hispanica L.). Eur J Clin Nutr. 2010;64(4):436-8. [ Links ]
23. Vuksan V, Choleva L, Jovanovski E, Jenkins AL, Au-Yeung F, Dias AG, et al. Comparison of flax (Linum usitatissimum) and Salba-chia (Salvia hispanica L.) seeds on postprandial glycemia and satiety in healthy individuals: a randomized, controlled, crossover study. Eur J Clin Nutr. 2017;71(2):234-8. [ Links ]
24. Vuksan V, Jenkins AL, Brissette C, Choleva L, Jovanovski E, Gibbs AL, et al. Salba-chia (Salvia hispanica L.) in the treatment of overweight and obese patients with type 2 diabetes: A double-blind randomized controlled trial. Nutr Metab Cardiovasc Dis. 2017;27(2):138-46. [ Links ]
25. Rank J, Nielsen MH. Allium cepa anaphase-telophase root tip chromosome aberration assay on N-methyl-N-nitrosourea, maleic hydrazide, sodium azide, and ethyl methanesulfonate. Mutat Res. 1997;390(1-2):121-7. [ Links ]
26. Leme DM, Marin-Morales MA. Allium cepa test in environmental monitoring: a review on its application. Mutat Res. 2009;682(1):71-81. [ Links ]
27. Glińska S, Bartczak M, Oleksiak S, Wolska A, Gabara B, Posmyk M, et al. Effects of anthocyanin-rich extract from red cabbage leaves on meristematic cells of Allium cepa L. roots treated with heavy metals. Ecotoxicol Environ Saf. 2007;68(3):343-50.
28. Sultan AÖ, Çelik TA. Genotoxic and Antimutagenic Effects of Capparis spinosa L. on the Allium cepa L. Root Tip Meristem Cells. Caryologia. 2009;62(2):114-23. [ Links ]
29. Akinboro A, Mohamed KB, Asmawi MZ, Sulaiman SF, Sofiman OA. Antioxidants in aqueous extract of Myristica fragrans (Houtt.) suppress mitosis and cyclophosphamide-induced chromosomal aberrations in Allium cepa L. cells. J Zhejiang Univ Sci B. 2011;12(11):915-22. [ Links ]
30. Sondhi N, Bhardwaj R, Kaur S, Kumar N, Singh B. Isolation of 24-epibrassinolide from leaves of Aegle marmelos and evaluation of its antigenotoxicity employing Allium cepa chromosomal aberration assay. Plant Growth Regul. 2007;54(3):217. [ Links ]
31. Carnier M, Silva FP, Miranda DA de, Hachul ACL, Silva Rischiteli AB, Pinto Neto NI, et al. Diet Supplemented with Chia Flour did not Modified the Inflammatory Process and Tumor Development in Wistar Rats Inoculated with Walker 256 Cells. Nutr Cancer. 2018;70(7):1007-16. [ Links ]
32. Marcinek K, Krejpcio Z. Chia seeds (Salvia hispanica): health promoting properties and therapeutic applications – a review. Rocz Panstw Zakl Hig. 2017;68(2):123-9. [ Links ]
33. Varela-Barca FNT, Agnez-Lima LF, de Medeiros SRB. Base excision repair pathway is involved in the repair of lesions generated by flavonoid-enriched fractions of pepper tree (Schinus terebinthifolius, Raddi) stem bark. Environ Mol Mutagen. 2007;48(8):672-81. [ Links ]
34. De Carvalho MCRD, Barca FNTV, Agnez-Lima LF, de Medeiros SRB. Evaluation of mutagenic activity in an extract of pepper tree stem bark (Schinus terebinthifolius Raddi). Environ Mol Mutagen. 2003;42(3):185-91. [ Links ]
35. Harwood M, Danielewska-Nikiel B, Borzelleca JF, Flamm GW, Williams GM, Lines TC. A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties. Food Chem Toxicol. 2007;45(11):2179-205. [ Links ]
36. Peh HY, Tan WSD, Liao W, Wong WSF. Vitamin E therapy beyond cancer: Tocopherol versus tocotrienol. Pharmacol Ther. 2016;162:152-69. [ Links ]
37. Sailo BL, Banik K, Padmavathi G, Javadi M, Bordoloi D, Kunnumakkara AB. Tocotrienols: The promising analogues of vitamin E for cancer therapeutics. Pharmacol Res. 2018;130:259-72. [ Links ]
Correspondencia:
Daniel López-Rojas
Arbitrado por pares
Recibido: 16/01/2019
Aprobado: 22/05/2019

