










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de la Sociedad Química del Perú
Print version ISSN 1810-634X
Rev. Soc. Quím. Perú vol.73 no.4 Lima Oct./dic. 2007
TRABAJOS ORIGINALES
Actividad antimicrobiana de una flavoproteína aislada del veneno de la serpiente peruana Bothrops atrox (jergón)
Antimicrobial activity of a flavoprotein isolated from the Bothrops atrox peruvian snake venom
Fanny Lazo1* ; Orestes Málaga1 ; Armando Yarlequé1 ; Ruperto Severino2 ; Susana Gutiérrez3
1 Lab. de Biología Molecular
2 Lab. de Zoología de Invertebrados
3 Lab. de Microbiología y Biotecnología Microbiana. Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos, Ciudad Universitaria, Av. Venezuela Cdra. 34, Lima 01.
* flazom@unmsm.edu.pe
RESUMEN
Se ha investigado la actividad de la L- aminoácido oxidasa (LAO), aislada previamente del veneno de la serpiente Bothrops atrox sobre cepas bacterianas Gram positivas y Gram negativas, asì como sobre promastigotes de Leishmania braziliensis braziliensis y epimastigotes de Trypanosoma cruzi. Adicionalmente se han ensayado las actividades edemática, hemorrágica y hemolítica de esta enzima.
El efecto antibacteriano, tanto del veneno crudo como de la enzima purificada, se evidenció utilizando la técnica de Grove sobre cultivos de Staphylococcus aureus, Streptococcus faecalis, Vibrio cholerae y Escherichia coli observándose que las bacterias Gram positivas son más susceptibles que las Gram negativas. Además, el veneno crudo de B. atrox y LAO presentaron un efecto in vitro, contra promastigotes de Leishmania braziliensis braziliensis con una CE50 de 7,82 y 1,33 µg/ml, respectivamente; en tanto que, sobre epimastigotes de Trypanosoma cruzi, el valor de CE50 fue de 7,95 µg/ml para el veneno crudo y 1,38 µg/ml para la enzima purificada. Adicionalmente, se ha observado que LAO no tiene actividad hemorrágica sobre piel de ratones albinos, ni hemolítica sobre eritrocitos humanos lavados. Sin embargo, produce edema, habiéndose calculado la DEM en 15,18 µg de proteína.
Palabras clave: Protozoario, bacteria, serpiente, enzima, veneno.
ABSTRACT
Antimicrobial activity on Gram positive and Gram negative bacteria was investigated using L- amino acid oxidase (LAO) from the venom of Bothrops atrox snake. Besides activity against promastigotes from Leishmania braziliensis braziliensis and epimastigotes from Trypanosoma cruzi were evaluated with either total venom and LAO. In addition, edemaforming, haemorragic and hemolytic effects of LAO were assayed.
The antibacterial effect of whole venom as well as purified enzyme was demonstrated by Grove's method on Staphylococcus aureus, Streptococcus faecalis, Vibrio cholerae and Escherichia coli grown cultures, being Gram positive, more sensible than Gram negative bacteria. In the same way B. atrox crude venom and its purified enzyme LAO, showed in vitro effect on Leishmania braziliensis braziliensis promastigotes (EC50 of 7,82 and 1,33 µg/ml respectively) and Trypanosoma cruzi epimastigotes (EC50 of 7,95 ug/ml for whole venom and 1,38 µg/ml for purified enzyme). LAO does not have neither haemorragic activity on mouse skin (20-22 g ) nor hemolytic activity on human red blood cells; however, LAO produces edema with a MED of 15,18 µg of protein.
Key words: Protozoa, bacteria, snake, enzyme, venom.
INTRODUCCIÓN
La L-aminoácido oxidasa (EC 1.4.3.2) es una flavoenzima responsable del color amarillo de los venenos de serpientes, que cataliza espicíficamente la desaminación oxidativa de un Laminoácido usado como substrato a un α-cetoácido con la producción de amoniaco y peróxido de hidrógeno. En los venenos de la familia viperidae, las L-aminoácido oxidasas se encuentran en una proporción promedio del 4% (p/p) del total de las proteína de la ponzoña (Iwanaga y Suzuki, 1979)1 por lo que estas muestras biológicas constituyen una buena fuente para su obtención (Mebs, 1970)2.
En los últimos años las LAOs de venenos de serpientes están siendo aisladas y purificadas para estudiar las interacciones que presentan durante los procesos de agregación plaquetaria (Takatsuka et al., 2001)3 e inducción de hemorragia y apoptosis (Torii et al., 2000)4. Asimismo, se ha encontrado que algunas enzimas de este tipo poseen propiedades antibacterianas, tal como lo demostró Stiles et al. (1991)5, investigando varias géneros de serpientes procedentes de América, Asia, África y Australia. Por su parte Yarlequé et al., (1997)6 determinaron la acción antibacteriana del veneno total y la enzima aislada de serpientes peruanas como Lachesis muta, Crotalus durissus terrificus. En cuanto a la acción antiprotozoaria, estudios realizados por Tempone et al., (2001)7 indicaron la capacidad letal de LAO sobre promastigotes de Leishmania spp.
En la presente investigación, se ha evaluado la acción antimicrobiana de LAO del veneno de B. atrox, serpiente que constituye un grave problema de salud en el Perú, habiéndose ensayado su actividad sobre bacterias Gram positivas y Gram negativas; así como también sobre promastigotes y epimastigotes de Leishmania braziliensis braziliensis y Trypanosoma cruzi, respectivamente.
MATERIALES Y MÉTODOS
Material biológico:
Veneno.- Se utilizó veneno crudo de especímenes adultos de la serpiente Bothrops atrox, procedentes de la zona de Pucallpa, Departamento de Ucayali, mantenidos en el serpentario Oswaldo Meneses del Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos. El veneno extraído por presión manual de las glándulas, fue liofilizado y conservado a –8 ºC.
Animales de experimentación.- Se usaron ratones albinos de ambos sexos (18 – 20 g).
Cantidad de proteína.- En todos los ensayos el contenido proteico del veneno crudo y de la proteína purificada se determinó, empleando el método de Lowry et al. (1951)8, usando como estándar de proteína, la albúmina sérica bovina y por el método de absorción ultravioleta a 280 nm (Warburg y Cristian, 1941)9.
Actividad antimicrobiana:
Actividad antibacteriana.- Se utilizaron cepas de Staphylococcus aureus ATCC 6538p, Vibrio cholerae Serotipo Inaba INS, Streptococcus faecalis INS CP 696 y Escherichia coli ATCC 51A266, usando el método de cilindro en placa (Grove y Randall, 1955)10 Los controles se hicieron colocando 10 µl de cloruro de sodio al 0,9% en lugar de las muestras en estudio. Asimismo, se hicieron pruebas con peroxidasa al 0,01% incubadas con la enzima purificada. Cada ensayo se realizó por triplicado.
Actividad antiprotozoaria.- Con respecto al efecto antiprotozoario, fue evaluado por ensayos de viabilidad celular sobre promastigotes de Leishmania braziliensis braziliensis y epimastigotes de Trypanosoma cruzi, según Tempone et al., (2000)7 y Fernández-Gómez et al. (1994)11.
Las muestras se incubaron a 25 C, extrayéndose alícuotas de 20 µl a intervalos de 4 horas para la observación microscópica y determinar la viabilidad de los protozoarios durante 24 horas, usando para el conteo una cámara de Neubauer. Los controles no contenían enzima y todas las pruebas fueron realizadas por triplicado.
También se hicieron ensayos colocando promastigotes de Leishmania braziliensis y epimastigotes de Trypanosoma cruzi, incubándolos con 20 µl de LAO (1 y 3 µg/ml) y peroxidasa 0,01% (20 µl) en un volumen final de 200 µl para observar el efecto antiprotozoario.
Los resultados obtenidos fueron tabulados empleando el método computarizado de Probits modificado por Castro De la Mata (1996)12, para establecer la concentración efectiva (CE50), medida que causa el 50% de mortalidad en el tiempo antes indicado.
Actividad biológica:
Efecto hemolítico.- Se empleó el método de Condrea et al. (1964)13, para lo cual se obtuvieron 9 ml de sangre venosa humana y se mezcló con 1 ml de citrato de sodio 3,8%. Luego de centrifugar a 3 500 rpm por 5 minutos, el plasma fue retirado y el paquete de glóbulos rojos se resuspendió y lavó 3 veces por centrifugación con 10 ml de cloruro de sodio 0,9% a pH 7,3, descartando en cada oportunidad el sobrenadante. Finalmente, los glóbulos rojos se resuspendieron en 10 ml de buffer isotónico (glicina 0,1M, ClNa 0,9% pH 7,3). La mezcla de reacción contenía 0,8 ml de buffer isotónico, 0,1 ml de glóbulos rojos y 0,1 ml de LAO (0,97 mg/ml) o del veneno crudo de B. atrox (5mg/ml) en buffer acetato de amonio 0,05M pH 7,4. Esta mezcla se incubó a 37 °C por 2 horas y la reacción se detuvo con 2 ml de cloruro de sodio 0,9% frío a pH 7,3 conteniendo EDTA 200 mM. Posteriormente se centrifugó a 3 500 rpm por 5 minutos para separar la hemoglobina libre a partir de eritrocitos lisados.
El grado de hemólisis se determinó por la cantidad de hemoglobina liberada y evaluada por espectrofotometrìa a una longitud de onda de 540 nm y fue expresada como porcentaje total de hemoglobina contenida en 0,1 ml de glóbulos rojos empacados y hemolizados totalmente con 5,9 ml de agua destilada
Actividad edemática.- Diferentes cantidades de LAO ( 2,5, 5, 10, 20 y 30 µg) disueltas en 50 µl de solución salina fueron inyectadas en el cojinete de la pata derecha de ratones albinos de 18 - 20 g de peso, empleándose para cada dosis grupos de cuatro ratones. En el cojinete de la pata izquierda se inyectaron 50 µl de solución salina (control).
Después de 3 horas, los ratones fueron sacrificados por dislocación cervical a fin de cortar y pesar las patas inyectadas. El edema fue expresado como el porcentaje del incremento del peso de la pata derecha con respecto a la pata izquierda. La dosis edemática mínima (DEM) es definida como la cantidad de toxina que produce un incremento de 30% en el peso (Yamakawa et al., 1976)14.
Actividad hemorrágica.- Se ensayó según el método de Kondo (1960)15, modificado por Lomonte et al. (1996)16. La cuantificación del efecto hemorrágico se logró inoculando por vía subcutánea a nivel abdominal, diferentes cantidades del veneno crudo (5, 10 y 15 µg) y de la enzima purificada (2,5, 5 y 10 µg), cada una en un volumen de 0,1 ml en ratones albinos; utilizándose para cada dosis grupos de 4 ratones. Dos horas después, los animales fueron sacrificados y se procedió a examinar las áreas hemorrágicas formadas. La dosis hemorrágica mínima (DHM), corresponde a la cantidad de proteína que produce un área hemorrágica de 10 mm de diámetro.
RESULTADOS Y DISCUSIÓN
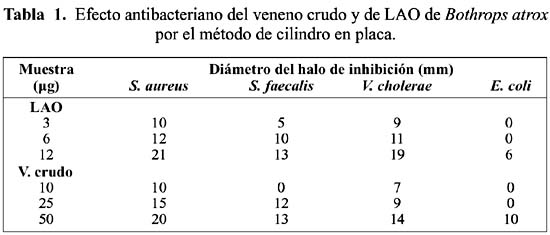
Efecto antibacteriano: La presencia de LAO en el rango de 3 a 12 µg, produjo un marcado halo de inhibición en el crecimiento de Staphylococcus aureus, Streptococcus faecalis (Gram positivas), así como en Vibrio cholerae bacteria Gram negativa (figura 1); en cambio, con Escherichia coli (Gram negativa) sólo se obtuvo halos de inhibición cuando se utilizaron 12 µg de LAO y 50 µg del veneno total, respectivamente (tabla 1). Asimismo, la preincubación de LAO con peroxidasa 0,01% ( 10 y 20 µl) dio lugar a una pérdida del halo de inhibición y por tanto de la actividad de LAO sobre S. aureus y V. cholerae.
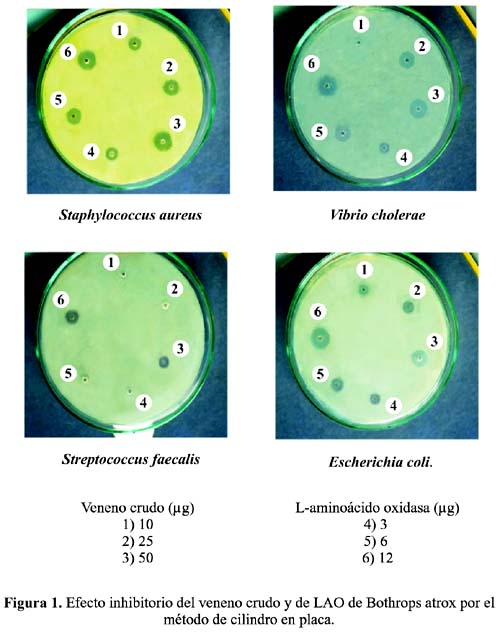
El efecto antibacteriano sobre S. faecalis puede atribuirse al hecho de que esta bacteria es catalasa negativa; en cambio S. aureus, pese a ser catalasa positiva, no produciría suficiente cantidad de catalasa durante su crecimiento para contrarrestar la producción de peróxido de hidrógeno formado por LAO y por lo tanto, el peróxido de hidrógeno actuaría como un agente bactericida, puesto que su acción fuertemente oxidante, en altas concentraciones, causa inestabilidad a nivel de membrana celular (Cisneros et al., 2006)17.
A la fecha no se conoce con precisión el mecanismo de acción de la enzima LAO implicada en la inhibición bacteriana, pero se sabe que su cofactor, el FAD, es reducido al reaccionar con los diferentes L-aminoácidos que se encuentran en el medio y que posteriormente la reacción enzimática da lugar a la reoxidación del FAD, con la liberación del peróxido de hidrógeno, al cual se le atribuye la acción antibacteriana, debido a su capacidad oxidante que lesiona membranas, proteínas y causa escisiones en los ácidos nucleicos.
Los resultados de esta investigación los podemos comparar con los datos reportados por Stiles et al., (1991)5, ya que al utilizar 6,25 µg de la enzima de C. adamanteus produjo un halo de inhibición de 10 mm sobre S. aureus mientras que, en el caso de LAO de B. atrox se utlizó 6 µg para producir un halo de 12 mm. De acuerdo al Manual Difco (1984) 30 µg de tetraciclina producen halos de inhibición mayores o iguales a 19 mm sobre bacterias Gram positivas o negativas y se considera que el microorganismo es sensible a la droga. En cambio, con la enzima purificada de B. atrox se encontró que con 12 µg sobre S. aureus y V. cholerae, se produjeron halos de inhibición de 21 y 19 mm, respectivamente.
Por otro lado, los estudios de Kornalik y Master (1964)18 han demostrado que el veneno de color amarillo obtenido de algunos especímenes de Vipera ammodytes contiene por lo menos 200 veces más LAO que aquellos de color blanco extraídos de otros ejemplares. Asimismo, dos venenos ligeramente amarillos como los de Naja melanoleuca y Naja naja oxiana fueron negativos, mientras que cinco venenos marcadamente amarillos de otras especies del género Naja fueron positivos en cuanto a la acción antibacteriana. Siendo la glándula venenosa una estructura modificada para producir diferentes proteínas, una de ellas podría estar encargada de la acción antibacteriana cuando la serpiente se alimenta, sobre todo si consideramos que estos ofidios ingieren mamíferos vivos y sin una previa masticación.
Efecto antiprotozoario.- Se observó in vitro la acción contra promastigotes de L. braziliensis, encontrándose una CE50 para el veneno crudo de 7,82 µg/ml y de 1,33 µg/ml para la enzima purificada (figura 2). Asimismo, contra epimastigotes de T. cruzi, se determinó la CE50 de 7,95 µg/ml para el veneno crudo y de 1,38 µg/ml para la enzima en estudio (figura 3). En cambio, cuando se incubaron promastigotes de L. braziliensis y epimastigotes de T. cruzi con LAO (1 a 3 µg/ml) más peroxidasa 0,01% (20 µl), se obtuvo un 100% de supervivencia, de los protozoarios analizados.
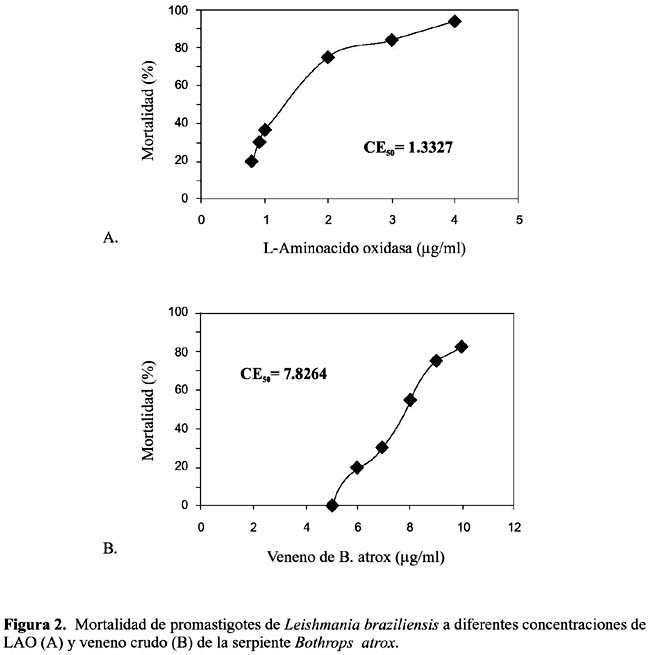
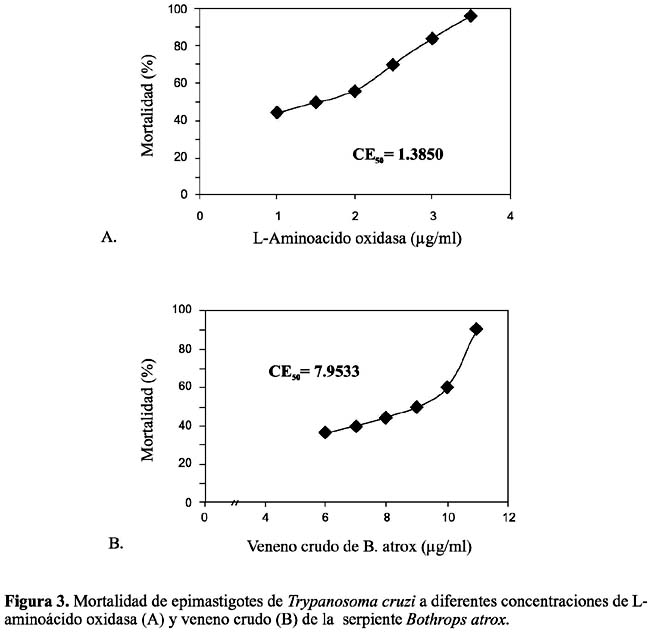
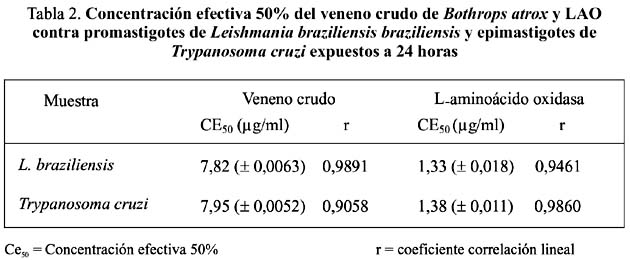
De estos resultados se puede deducir que la actividad antileishmánica de LAO es seis veces más alta que la del veneno total; en tanto que para T. cruzi la acción de LAO es 4, 8 veces mayor (tabla 2). Debido a los pocos estudios realizados hasta la fecha en el análisis antiprotozoario de enzimas purificadas de venenos ofìdicos, esta investigación es pionera para nuestro país y muestra que una proteína como LAO puede interferir no sólo el crecimiento y supervivencia de algunas bacterias patógenas sino también de protozoarios flagelados causantes de dolencias como Leishmaniasis y la terrible enfermedad de Chagas Tempone et al., (2001)7; Fernández-Gómez et al. (1994)11.
Tal vez, una explicación para comprender el efecto anti leishmánico de LAO sería que el estrés oxidativo producido por el peróxido de hidrógeno podría activar el shock proteico descrito para Leishmania sp, ya que se induciría la actividad proteolítica dentro de la célula y también se afectaría la función mitocondrial debido al incremento de las concentraciones de calcio. Es importante resaltar que Tempone et al. (2001)7 mostraron que no hay evidencia de otras proteínas en el veneno de B. moojeni con actividad antileishmánica. Es también interesante notar que la incubación de LAO con amastigotes de Leishmania no los afecta, lo que sugiere que en este estadio dichos parásitos tienen un eficiente sistema de protección contra radicales libres y peróxido de hidrógeno; las enzimas catalasa, superóxido dismutasa y otras, actuarían en el mecanismo de defensa antioxidante. En cambio, los promastigotes son deficientes en catalasa y glutation peroxidasa y pueden ser eliminados en un 80 - 95% por el peróxido de hidrógeno producido por los macrófagos durante la infección
3. Actividad edemática.- La enzima purificada del veneno de B. atrox fue capaz de inducir edema en el cojinete plantar de ratones albinos. Los incrementos en peso obtenidos con 5, 10, 15 y 20 µg fueron 10,3, 14,7, 27,6 y 43,13%, respectivamente, determinándose una DEM de 15,18 µg de proteína (figura 4). Asimismo se ha demostrado que LAO de B. atrox carece de las actividades hemorrágica y hemolítica, las cuales sí están presentes en el veneno total.
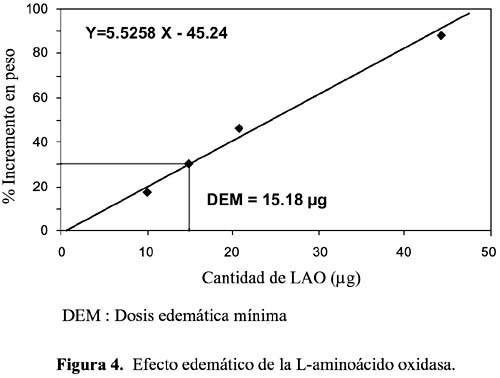
Muy pocos estudios se han realizado para determinar la naturaleza exacta del mecanismo del edema causado por LAO, aunque en algunos casos se ha calculado la dosis edemática mínima (DEM); así, por ejemplo, para Ophiophagus hannah cobra real, la DEM es 6,1-6,8 µg/pata en ratones, 8 µg/pata de ratón para Trimeresurus mucrosquamatus y para Eristocophis macmahoni 4,8 µg/ml (Yamakawa et al. 1976)14.
En nuestro caso, para la enzima purificada de Bothrops atrox, hemos encontrado una DEM de 15,18 µg de proteína (figura 4), lo que nos indica que LAO de este veneno tiene una moderada acción edemática equivalente a un 30% del máximo valor encontrado para una enzima de este tipo procedente de un veneno ofídico. Estudios previos revelaron que el edema inducido por venenos de serpientes es debida a la acción directa de alguna de sus toxinas o son mediadas por la liberación de autocoides como histamina, 5-hidroxitriptamina y sustancias de baja reactividad, como prostaglandinas, serotoninas y kininas, (Ali et al., 1999)19.
Asimismo, se ha reportado que existen otros inductores de edema, como ciertas proteasas y fosfolipasas ofídicas.
Por otro lado, se tienen evidencias que la actividad de LAO de O. hannah, que induce la formación de edema, no fue inhibida por difenhidramina o dexametasona, que son drogas antihistamínicas; sin embargo, preadministrando glutatión a los ratones se inhibe la actividad de la enzima, sugiriendo que el peróxido de hidrógeno liberado por acción desaminativa de la proteína está involucrado en la formación del edema, debido a que reduce a la flavina, lo que permitiría asumir que el FAD está directamente involucrado en el mecanismo catalítico (Tan y Ponnudurai, 1991)20. Otro posible camino para la acción inhibidora del glutatión sobre LAO, es su capacidad para escindir enlaces disulfuros presentes en la enzima, debido a que la proteína posee un grupo sulfihidrilo.
Otro aspecto muy interesante de resaltar es que si algunas L-aminoácido oxidasas ofídicas son capaces de inducir edema, tal como ocurre con la de B. atrox, esto sería una evidencia de su activa participación en el proceso inflamatorio posterior a la mordedura, por ello un estudio exhaustivo sobre el modo de bloquear a la enzima permitiría desarrollar métodos terapéuticos de gran utilidad en el control de este mecanismo.
AGRADECIMIENTOS
Los autores de este trabajo expresan su agradecimiento a las biólogas Gloria Sáez y Rosa Martínez de los laboratorios de Microbiología y Parasitología Ambientales de la UNFV y de Parasitología de Fauna Silvestre de la UNMSM, respectivamente, por habernos proporcionado las cepas de protozoarios analizados.
REFERENCIAS BIBLIOGRÁFICAS
1. Iwanaga S, Suzuki T. Enzymes in snake venoms. In: Lee CY, editor. Handbook of Experimental Pharmacology (Snake Venoms). Berlin: Springer-Verlag; 1979,52: 75-84.
2. Mebs D. A comparative study of enzyme activities in snake venoms. International J Biochem. 1970; 1: 335-342.
3. Takatsuka H, Sakurai Y, Yoshioka A, Kokubo T, Usami y, Suzuki M, Matsui T, Titani K, Yagi H, Matsumoto M, Fujimura Y. Molecular characterization of L- amino acid oxidase from Agkistrodon halys blomhoffii with special reference to platelet aggregation. Biochim Biophys Acta 2001; 1544: 267-277.
4. Torii S., Naito M., Tsuruo T. Apoxin I, a novel apoptosis-inducing factor with L- amino acid oxidase activity purified from Western diamondback rattlesnake venom. J Biol Chem 1997; 272 (14): 9539-9542.
5. Stiles B.G., Sexton F.W., Weinstein S.A. Antibacterial effects of different snake venoms: purification and characterization of antibacterial proteins from Pseudechis australis (Australia King brown or mulga snake) venom. Toxicon 1991; 29: 1129-1141.
6. Yarlequé A., Cárdenas J., Escobar E., Gutierrez S. Some biochemical properties and antibacterial action of a L- amino acid oxidase from Peruvian snake venoms. Toxicon 1997; 35(4): 489.
7. Tempone A.G., Andrade H.F. JR, Spencer P.J., Lourenco C.O., Rogero Jr., Nascimento N. Bothrops moojeni venom kills Leishmania spp. With hydrogen peroxide generated by its L- amino acid oxidase. Biochemical and Biophysical research Communications 2001; 280(3): 620-624.
8. Lowry O.H., Rosebrough N.J., Farr A.L., Randall R. Protein measurement with the folin phenol reagent. J Biol Chem 1951; 193: 265-275.
9. Warburg O., Christian W. Isolierung and cristallisation del Garungs ferments enolase. Biochemische Zertschrift 1941; 31: 384-421.
10. Grove d., Randall W. Assay of antibiotics. 1st ed. New York: Medical Encyclopedia Inc; 1955. p. 78-86.
11. Fernández-Gómez R., Zerrouk H., Sebti F., Loyens M., Benslimane A., Ouaissi A. Growth inhibition of Trypanosoma cruzi and Leishmania donovani infantum by different snake venoms: Preliminary identification of proteins from Cerastes cerastes venom which interact with the parasites. Toxicon 1994; 32(8): 875-882.
12. Castro de la Mata R. Manual del programa de estadística para ensayos biológicos. Servicio de control de calidad. UPCH. Lima Perú; 1996.
13. Condrea E., De Vries A., Mager J. Hemolysis and splitting of human erytrocyte phospholipids by snake venom. Biochim Biophys Acta 1964; 84:60.
14. Yamakawa M., Nosaki M., Hokama Z. Fractionation of sakishima-habu (Trimeresurus elegans) venom, and lethal, hemorrhagic and edema-forming activities of the fractions. In: Ohsaka A, Hayashi K, Sawai Y, editors. Animal, Plant and Microbial Toxins. Vol1, Biochemistry, New York: Plenum Press; 1976.
15. Kondo H., Kondo S., Ikesawa M., Murata R., Ohsaka A. Studies on the cuantitative method for determination of hemorrhagic activity of Habu snake venom. J Med Sci Biol 1960; 13:43.
16. Lomonte B., Leon G., Hanson L. Similar effectiveness of Fab and F(ab') antivenom in the 2 neutralization of hemorrhagic activity of Vipera berus snake venom in mice. Toxicon 1996; 34(10): 1197-1202.
17. Cisneros Y., Lazo F., Gutierrez S., Yarlequé A. Caracterìsticas bioquímicas de una proteína antibacteriana aislada del veneno de Lachesis muta shushupe. Rev Soc Quim Perú 2006; 72(4):187-196.
18. Kornalik F., Master R.W.P. A comparative examination of yellow and white venoms of Vipera ammodytes. Toxicon 1964; 2: 109-115.
19. Ali S.A., Stoeva S., Abbasi A., Alam J.M., Kayed R., Faigle M., Neumeister B., Voelter W. Isolation structural and functional characterization of na apoptosis-inducing L- amino acid oxidase from leaf- nosed viper (Eristocophis macmahoni) snake venom. Arch Biochem Biophys 2000; 384: 216-226.
20. Tan N.H., Ponnudurai G. A comparative study of the biological properties of some sea snake venoms. Comp Biochem Physiol 1991; B99: 351-354.
