










Services on Demand
Journal

Article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkScientia Agropecuaria
Print version ISSN 2077-9917
Scientia Agropecuaria vol.5 no.3 Trujillo 2014
http://dx.doi.org/10.17268/sci.agropecu.2014.03.03
ARTÍCULOS ORIGINALES
Densidade e distribuição estomática em mandioca Manihot esculenta Crantz cultivar IAC 576-70
Density and stomata distribution in cassava Manihot esculenta Crantz IAC 576-70
Eli Carlos de Oliveira*; Édison Miglioranza
Universidade Estadual de Londrina – UEL. Londrina-PR.
Resumo
Em plantas de mandioca a variação nas características estomáticas, como a densidade, o diâmetro e a funcionalidade sofrem alterações por conta da frequência e distribuição dos estômatos nas diferentes regiões do limbo foliar. Este estudo teve como objetivo determinar a densidade, diâmetros e a distribuição dos estômatos em diferentes regiões da folha de mandioca da cultivar IAC 576-70. Foram analisadas as regiões apical, mediana e basal de folhas completamente expandidas. O delineamento foi inteiramente casualizado com três regiões amostradas e vinte repetições, sendo cada região da folha observada em cinco campos microscópicos. Os dados foram submetidos à análise de variância e suas médias comparadas pelo teste de Tukey. Os parâmetros avaliados foram dados referentes ao número, diâmetro polar e equatorial dos estômatos e cálculos da densidade e a funcionalidade estomática. Nao há diferença entre a densidade estomática, diâmetro equatorial e funcionalidade dos estômatos nas diferentes regiões da folha de mandioca da cultivar IAC 576-70, enquanto que o diâmetro polar dos estômatos é menor na região apical e mediana das folhas.
Palavras-chave: diâmetro polar, diâmetro equatorial, funcionalidade estomática.
Abstract
In cassava the variation in stomata characteristics, such as density, polar diameter and functionality undergo changes due to the stomata frequency distributed in different regions of the leaf blade. This study aimed to determine the density, diameters and distribution of stomata in different regions of cassava leaf. We analyzed the apices, median and basal expanded leaves of IAC 576-70. The experimental design was completely randomized with three regions sampled twenty replications, each leaf region observed in five microscopic fields. Data were subjected to analysis of variance and the averages compared by Tukey test. We evaluated data on the number, polar and equatorial diameter of the stomata and calculations of stomata density and functionality. There is no difference between stomata density, equatorial diameter and functionality of stomata in different regions of cassava leaf cultivar IAC 576-70, while the polar diameter of the stomata is smaller in the apical and median regions of leaves.
Keywords: polar diameter, equatorial diameter, stomata functionality.
1. Introdução
Pertencente à família Euphorbiaceae, a mandioca (Manihot esculenta Crantz), é uma planta cultivada em várias regiões do mundo, sob diferentes condições de temperaturas, precipitação e tipo de solo, além de possuir vasta variabilidade genética (Ascenio, 1996).
Embora os aspectos anatômicos da mandioca sejam ainda pouco estudados, há importância no conhecimento dos mesmos devido às modificações na anatomia foliar terem sido detectadas em diferentes cultivares de mandioca, promovendo diferenças na taxa fotossintética por modificações na difusão do CO2 (El-Sharkawy, 2006). Diferentes cultivares de mandioca evidenciam a plasticidade para a anatomia foliar, exibindo diferenças nas características estomáticas, como a densidade, que podem demonstrar cultivares em potencial para a resistência à seca (Cerqueira, 1992).
A anatomia vegetal pode indicar características que atribuem tolerância a diferentes condições ambientais para as plantas cultivadas, como tolerância à seca (Grisi et al., 2008; Batista et al., 2010) como a qualidade e intensidade da radiação que incidem sobre as folhas (Santiago, 2001) ou mesmo estresses causados por alagamentos (Souza, 2007; Souza et al., 2010) promovendo modificações na espessura do mesofilo, na espessura da epiderme, na densidade e tamanho de estômatos e demais características estomáticas que conferem tolerância a essas condições. A variação nas características estomáticas, como densidade, diâmetro polar e funcionalidade também sofrem alterações por conta da frequência estomática distribuídos em diferentes regiões do limbo foliar (Cock et al., 1987).
Diante do exposto o presente estudo teve como objetivo determinar a densidade, os diâmetros e a distribuição dos estômatos em diferentes regiões da folha de mandioca da cultivar IAC 576-70.
2. Material e métodos
A pesquisa foi realizada na Universidade Estadual de Londrina (UEL), Estado do Paraná, localizada a, latitude 23°23 S, longitude 51°11 O e 566 m de altitude. A pluviosidade média anual é de 1400 mm; a temperatura média do ar é de 25,5°C, com pequenas diferenças entre as médias mensais, cerca de 2°C, a umidade relativa do ar oscila entre 60% na estação seca e 75% na estação chuvosa.
Para o estudo foi utilizado a cultivar IAC 576-70 que representa praticamente a totalidade das áreas destinadas ao comércio de mandioca de mesa na região da pesquisa. Cultivar desenvolvido pelo IAC a partir do cruzamento entre SRT 797 Ouro do Vale e IAC 14-18, é superior aos demais em cultivo, por sua produtividade, aspecto das raízes, qualidades culinárias e sensoriais, e considerável resistência à bacteriose (Lorenzi et al., 1984). Foram analisadas as regiões apical, mediana e basal de folhas completamente expandidas em três plantas, as quais foram plantadas no dia 27 de junho de 2012 no solo tipo Latosolo vermelho escuro de textura média. A adubação de base foi realizada com 560 kg ha-1 do adubo formulado 08-28-16 (NPK). Os lóbulos foliares foram coletados no dia 30 de abril de 2013 das 09:00 as 11:30 horas, onde não houve registro de precipitação, enquanto que a temperatura mínima e máxima registravam 14 e 28 °C respectivamente. As amostras de folhas foram fixadas em solução, contendo 5% de formol, 90% de álcool etílico a 70°GL e 5% de ácido acético (F.A.A), por 24 horas à temperatura ambiente. Preparouse o material foliar segundo a metodologia proposta por Johansen (1940), para visualização e caracterização do tipo de estômato através de observação por microscópio eletrônico de varredura (MEV). A observação da densidade e da distribuição dos estômatos foi realizada empregandose o método de réplica de North (1956), onde uma impressão da superfície foliar foi obtida com acetato de celulose com uma fita adesiva transparente em uma área de aproximadamente de 1,0 cm2. Seções da película de acetato formada foram removidas e montadas em lâminas para serem avaliadas ao microscópio. Para as observações, utilizou um microscópio ótico, ocular de 15x e objetiva de 40x, o que proporcionou aumento de 600x e campo real visível de 0,39mm2. As imagens utilizadas para análise foram capturadas por meio de um sistema de captura (Carl Zeiss, Germany) composto de: microscópio AxioPhot I, equipado com câmera AxioCam ICc3 e software Bel View. O processamento e análise das imagens foram realizados através do software de domínio público ImageJ 1.43a, versão 64, com a medição de cinco campos por repetição para cada variável analisada. Para as avaliações, diâmetro polar dos estômatos (DP) e diâmetro equatorial dos estômatos (DQ), preparouse laminas com safranina, sendo observadas em câmara clara, segundo técnica de Labouriau (Labouriau et al., 1961). Foram calculados a densidade estomática (DE - número de estômatos por unidade de área) e a funcionalidade estomática (FUN – considerada como a relação diâmetro polar/diâmetro equatorial dos estômatos) segundo Castro et al. (2009).
Para efeito de análise estatística, estudouse a densidade dos estômatos na face abaxial e a frequência dos estômatos nas regiões da base, meio e ápice dos lóbulos foliares. O delineamento foi inteiramente casualizado com três regiões amostradas e vinte repetições, sendo cada região da folha observada em cinco campos microscópicos. Os dados foram submetidos à análise de variância e suas médias comparadas pelo teste de Tukey (p < 0,05). Como os dados apresentaram distribuição normal (Shapiro-Wilk) nenhuma transformação foi utilizada antes de realizar a análise de variância. Para análise estatística foi utilizado o software o Sisvar (Ferreira, 2004).
3. Resultados e discussão
A análise conjunta dos dados foi realizada para as três regiões do lóbulo foliar. Observouse efeito significativo apenas para o parâmetro diâmetro polar (DP) (p < 0,05) (Tabela 1).
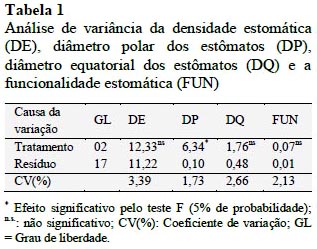
Como pode ser observado pela Tabela 1, não houve diferenças significativas em relação à posição da folha verificada, para a densidade de estômatos (DE), diâmetro equatorial (DQ) e funcionalidade (FUN) (p > 0,05).
Os estômatos foram classificados como do tipo paracíticos, estando distribuídos de forma ondular na face abaxial do lóbulo foliar. Foram observadas através de MEV, na face abaxial do lóbulo foliar, células formando papilas com uma cúpula mais ou menos cônica envolta do estômato (Figura 1).

Foi verificada a presença destas papilas por toda a epiderme da face abaxial, porém, ao lado das nervuras primárias e secundárias apresentam um formato mais arredondado com cúpulas de menor projeção (Figura 2).

Trabalhos como os de Theobald et al. (1979), Bone et al. (1985), Mantovani e Vieira (1997) relatam a presença das papilas na face abaxial da epiderme nas duas faces das folhas em plantas da família Euphorbiaceae. Características semelhantes foram encontradas na epiderme da face adaxial de Manihot caerulescens, descritas por Mendonça (1983). Em Manihot glaziovii, Mendonça (1992) observou a ocorrência de células retangulares, com as paredes periclinais externas frequentemente pontiagudas, assemelhandose, às vezes, com papilas.
As presenças destas papilas na epiderme contribuem para a melhoria na recepção de luz em plantas que crescem debaixo do dossel de florestas tropicais (Theobald et al., 1979; Bone et al., 1985; Mantovani e Vieira 1997). Considerando que M. esculenta é uma planta cultivada em pleno sol, a presença dessas papilas na face abaxial parece ter a função principal de proteger os estômatos, já que se distribuem como uma coroa ao redor deles. No entanto, a comprovação da função das papilas como proteção estomática ou mesmo como lentes que poderia convergir às luzes para os tecidos da clorofila, necessita de estudos mais aprofundados.
Os dados referentes à densidade estomática (DE), diâmetro polar (DP), diâmetro equatorial (DQ) e funcionalidade estomática (FUN) estão apresentados na tabela 2.
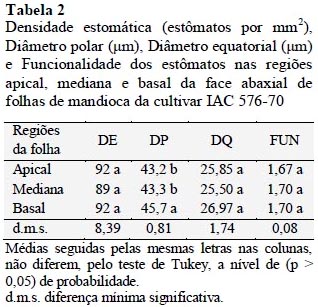
A densidade estomática não diferenciou significativamente entre as diferentes posições da folha, ao contrário do esperado. Muitos estudos apontam uma correlação positiva entre a posição da folha e a densidade estomática (Schonherr e Bukovac, 1972; Rodella et al., 1982; Mcwhorter et al., 1993; Heichel, 1999).
Os estômatos estão associados diretamente com a capacidade fotossintética, e, uma alteração na sua quantidade afeta diretamente a condutância estomática, ou seja, quanto maior a densidade estomática, maior a absorção de CO2 (Abrans et al., 1992; Evans, 1999). O tamanho, a posição e o controle de abertura dos estômatos também irão afetar na absorção de CO2 (Boeger e Wisniewski, 2003).
Verificouse para os valores médios de DP que os maiores valores estão na região basal da folha, apresentando diferença significativa entre a região apical e mediana que não diferiram entre si, apresentando os menos valores para esta variável (p < 0,05). Ainda é possível afirmar que a redução no tamanho dos estômatos é um evento reconhecidamente importante na regulação das trocas gasosas, uma vez que folhas com estômatos menores apresentam maior eficiência no uso da água por apresentarem um menor tamanho dos poros estomáticos, condicionando assim uma menor perda de água por transpiração (Boeger e Wisniewski, 2003).
Segundo Kerbauy (2004) a redução no tamanho da abertura estomática apresenta maior efeito sobre a difusão de água do que sobre a difusão de CO2, podendo, desse modo, manter o influxo de CO2 necessário à fotossíntese e uma menor perda de água por transpiração. Neste mesmo sentido, o aumento na densidade estomática, acoplada à diminuição no tamanho dos estômatos. Variações no comportamento estomático, em relação ao seu tamanho, é uma característica muito variável em plantas em função do ambiente (Melo et al., 2004), da constituição genética da espécie (Alves et al., 2001; Camargo e Marenco, 2011), e frequentemente em plantas submetidas a diferentes estresses (Castro et al., 2005).
Com relação ao DQ e FUN não foram observados diferenças significativas entre as posições das folhas amostradas da cultivar de mandioca IAC 576-70 (p > 0,05).
As características avaliadas nesta pesquisa acima mostram a plasticidade morfológica de folha de mandioca. Nas condições estabelecidas pela amostragem abordada na pesquisa permitem inferir que a cultivar IAC 576-70 possui capacidade de adaptarse a diferentes habitats, de acordo com as confirmações de Mendonça (1983).
Porém, o que se percebe é que plantas de mandioca, mais particularmente as características das folhas, órgão responsável pela síntese de carboidratos e principal responsável pela tuberização da raiz, apresenta características importantes na distribuição e dimensões de seus estômatos em função do genótipo, além de uma anatomia foliar própria e específica, como a presença de papilas na face abaxial da epiderme, o que pode estar contribuindo de forma protetiva aos estômatos, e com isto mantendo que os mesmos permaneçam por maior tempo abertos mesmo em ambientes com déficit hídrico.
4. Conclusões
De acordo com a pesquisa e os resultados obtidos, é possível concluir que: não há diferença entre a densidade estomática, diâmetro equatorial e funcionalidade dos estômatos nas diferentes regiões da folha de mandioca da cultivar IAC 576-70. E que o diâmetro polar dos estômatos é menor na região apical e mediana das folhas da cultivar IAC 576-70.
5. Agradecimentos
O primeiro autor agradece a Fundação Araucária pela bolsa concedida.
6. Referências bibliográficas
Abrans, M.C.; Kloeppel, B.D.; Kubiske, M.E. 1992. Ecophysiological and morphological responses to shade and drought in two contrasting ecotypes of Prunus serotina. Tree physiology 10: 343-355. [ Links ]
Alves, E.S.; Giusti, P.M.; Domingos, M. 2001. Anatomic studies on Trandescantia hibrid clone 4430 leaves: changes caused by urban air pollution. Revista Brasileira de Botânica 24: 561-566. [ Links ]
Ascenio, J. 1996. Algunos aspectos relacionados con la fisiología de la planta de yuca. En: Montaldo A (ed.). La yuca frente al hambre del mundo tropical. Universidad Central de Venezuela, Venezuela. [ Links ]
Batista, L.A., Guimarães, R.J.; Pereira, F.J.; Carvalho, G.R.; Castro, E.M. 2010. Anatomia foliar e potencial hídrico na tolerância de cultivares de café ao estresse hídrico. Revista Ciência Agronômica 41: 475-481. [ Links ]
Boeger, M.R.T.; Wisniewski, C. 2003. Comparação da morfologia foliar de espécies arbóreas de três estádios sucessionais distintos de Floresta Ombrófila Densa (Floresta Atlântica) no sul do Brasil. Revista Brasileira de Botânica 26: 61-72. [ Links ]
Bone, R.A.; Lee, D.W.; Norman, J.M. 1985. Epidermal cells functioning as lenses in leaves of tropical rainforest shade plants. App. Opt. 24: 1408-12. [ Links ]
Camargo, M.A.B.; Marenco, R.A. 2011. Density, size and distribution of stomata in 35 rainforest tree species in Central Amazonia. Acta Amazonica 41: 205-212. [ Links ]
Castro, E.M.; Pinto, J.E.B.P.; Melo, H.C.; Soares, A.M.; Alvarenga, A.A.; Lima, E.C. 2005. Aspectos anatômicos e fisiológicos de plantas de guaco submetidas a diferentes fotoperíodos. Horticultura Brasileira 23: 846-850. [ Links ]
Castro, E.M.; Pereira, F.J.; Paiva, R. 2009. Histologia Vegetal: estrutura e função dos órgãos vegetativos. UFLA, Lavras. [ Links ]
Cerqueira, Y.M. 1992. Efeito da deficiência de água na anatomia foliar de cultivares de mandioca Manihot esculenta Crantz. I-Densidade estomática. Sitientibus 10: 103-115. [ Links ]
Cock, J.H.; Riano, N.M.; El-Sharkawy, M.A; Lopes, F.Y.; Bastidas, G. 1987. C3-C4 intermediate photosynthetic characteristics of cassava (Manihot esculenta Crantz) - II. Initial products of 14 CO2 fixation. Photosynthesis Research 12: 237-241. [ Links ]
El-sharkawy, M.A. 2006. International research on cassava photosynthesis, productivity, ecophysiology, and responses to environmental stresses in the tropics. Photosynthetica 44: 481-512. [ Links ]
Evans, J.R. 1999. Leaf anatomy enables more equal access to light and CO2 between chloroplasts. New Phytologist 143: 93-104. [ Links ]
Ferreira, D.F. 2004. SISVAR - versão 4.6. DEX/UFLA, Lavras. [ Links ]
Grisi, F.A.; Alves, J.D.; Castro, E.M.; Oiveira, C.; Biagiotti, G.; Melo, L.A. 2008. Avaliações anatômicas foliares em mudas de café 'Catuaí' e 'Siriema' submetidas ao estresse hídrico. Ciência e Agrotecnologia 32: 1730-1736. [ Links ]
Heichel, G.H.H. 1999. Genetic control of epidermal cell and stomatal frequency in maize. Crop Science 11: 830–832. [ Links ]
Johansen, B.A. 1940. Plant microtechnique. McGraw-Hill Book. New York. [ Links ]
Kerbauy, G.B. 2004. Fisiologia Vegetal. Editora Guanabara Koogan, São Paulo. [ Links ]
Labouriau, L.G.; Oliveira, J.G.; Salgado-Labouriau, M.L. 1961. Transpiração de Schizolobium parahyba(Vell.) Toledo I. Comportamento na estação chuvosa, nas condições de Caeté, Minas Gerais, Brasil. An. Acad. Bras. Ci. 35: 237-257. [ Links ]
Lorenzi, J.O.; Elias, R.; Valle, T.L.; Peressin, V.A.; Scarpare, J.A. filho. 1984. Variedade de mandioca de mesa IAC 576-70. Campinas: Coordenadoria de Assistência Técnica Integral CATI. Comunicado técnico 80: 1-4. [ Links ]
Mantovani, A.; Vieira, R.C. 1997. Leaf surface of two understorey shrubs Rudgea decipiens Müll. Ar. and Rudegea macrophylla Bent. (Rubiaceae). Rodriguesia 45: 7-13. [ Links ]
Melo, H.C.; Castro, E.M.; Ferrer, J.L.R.; Alves, J.D.; Fries, D.D.; Melo, L.A.; Magalhães, P.C. 2004. Influência da aplicação de cálcio e alagamento do solo sobre características anatômicas das folhas de milho (Zea mays L.) "Saracura" BRS-4154. Revista Brasileira de Milho e Sorgo 3: 333-342. [ Links ]
Mendonça, M.S. 1983. Estudo de plantas laticíferas. I. Aspectos anatômicos e distribuição de vasos laticíferos em Manihot caerulencens Pohl. Acta Amazônica 13: 501-17. [ Links ]
Mendonça, M.S. 1992. Estudo de plantas laticíferas. II Aspectos anatômicos e distribuição de vasos laticíferos em Manihot glaziovii Muller Arg. Acta Amazônica 22: 309-21. [ Links ]
Mcwhorter, C.G.; Ouzts, C.; Paul, R.N. 1993. Micromorphology of johnsongrass (Sorghum halapense) leaves. Weed Science 41: 583-589. [ Links ]
North, C,A. 1956. Technique for measuring structural features of plant epidermis using cellulose acetate films. Nature 178: 1186–1187. [ Links ]
Rodella, R.A; Ishiy; C.M.; Maiomoni, R.C.S.; Amaral Junior, A. 1982. Número e distribuição dos estômatos em folhas de duas espécies de Brachiária, Revista Poliagro 4: 1–10. [ Links ]
Santiago E.J.A. 2001. Aspectos da anatomia foliar da pimenta-longa (Piper hispidinervium C. DC.) sob diferentes condições de luminosidade. Ciência e Agrotecnologia 25: 1035-1042. [ Links ]
Schooner, J.; Bukovac, M.J. 1972. Penetration of stomata by liquids: dependence on surface tension, wettability, and stomatal morphology. Plant Physiology 49: 813-819. [ Links ]
Souza, G.S. 2007. Estrutura foliar e de cloroplastídeos em Mikania laevigata Shultz Bip. ex Baker em diferentes condições de qualidade de luz. Revista Brasileira de Biociências 05: 78-80. [ Links ]
Souza, T.C.; Magalhaes, P.C.; Pereira, F.J.; Castro, E.M.; Silva Júnior, J.M.; Parentoni, S.N. 2010. Leaf plasticity in sucessive selection cycles of "Saracura" maize in response to soil flooding. Pesquisa Agropecuária Brasileira 45: 16-24. [ Links ]
Theobald, W.L.; Krahulik, J.L.; Rollins, R.C. 1979. Tricome Description and classification. In: METCALFE, C.R., CHALK, L. Anatomy of the dicotyledons. 2.Ed. Oxford: Claredon Press. [ Links ]
* Autor para correspondencia
E-mail: elioliveira.agro@gmail.com (E. de Oliveira).
Recibido 18 julio 2014.
Aceptado 21 septiembre 2014.

