










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkScientia Agropecuaria
versión impresa ISSN 2077-9917
Scientia Agropecuaria vol.9 no.3 Trujillo jul./set. 2018
http://dx.doi.org/10.17268/sci.agropecu.2018.03.11
Artículos originales
Comportamiento de parámetros biométricos de clones para la obtención de papa baby con pulpa pigmentada
Behavior of biometric parameters clones for the obtaining of potato baby with pigmented flesh
Roberto Tirado-Lara1,*; Roberto Hugo Tirado-Malaver2
1 Universidad Nacional Pedro Ruiz Gallo, Calle Juan XXIII, Lambayeque, Perú.
2 Universidad Nacional Faustino Sánchez Carrión, Pje. Mercedes Indacochea 609, Huacho, Perú.
Resumen
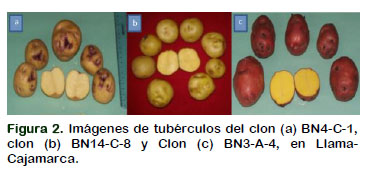
En el Perú las papas con pulpa pigmentada tienen un gran valor científico y biológico, son cultivadas y preservadas en áreas muy localizadas por pequeños agricultores. Se analizó el comportamiento de clones de papa con pulpa pigmentada en Llama, Cajamarca, con el objetivo de obtener variedades de alta producción de minitubérculos denominadas como "papa baby". Para ello se seleccionaron veinte clones provenientes de las hibridaciones entre cultivares nativos de Cajamarca con ecotipos nativos. Respecto al número de tubérculos por planta, los mejores clones fueron: BN4-C-1 de color de piel y pulpa crema, BN14-C-8 de piel y pulpa crema, BN3-A-4 de piel roja y pulpa amarilla y BN14-V-2 color de piel y pulpa violeta, con 90,73, 42,9, 36,87 y 31,14 tubérculos por planta. Además, estos clones presentaron el mayor número de tubérculos no comerciales por planta con 57,50, 13,87, 13,07 y 10,07, número de tallos por planta con 3,33, 3,43, 2,40 y 2,93, respectivamente. Siendo estos clones las que obtuvieron mayor número de minitubérculos para la comercialización y consumo humano como papa baby.
Palabras clave: comercialización; cultivares nativos; ecotipos nativos; regresión; variedades.
Abstract
In Peru, potatoes with pigmented pulp have a great scientific and biological value, are cultivated and preserved in areas very localized by small farmers. The behavior of potato clones with pigmented pulp was analyzed in Llama, Cajamarca, in order to obtain high production varieties of minitubers called "baby potatoes". For this, twenty clones were selected from the hybridizations between native cultivars of Cajamarca with native ecotypes. Regarding the number of tubers per plant, the best clones were: BN4-C-1 skin color and cream pulp, BN14-C-8 skin and cream pulp, BN3-A-4 red skin and yellow flesh and BN14 -V-2 skin color and violet pulp, with 90.73, 42.9, 36.87 and 31.14 tubers per plant. In addition, these clones had the highest number of non-commercial tubers per plant with 57.50, 13.87, 13.07 and 10.07, number of stems per plant with 3.33, 3.43, 2.40 and 2.93, respectively. Being these clones those that obtained greater number of minitubers for the commercialization and human consumption like baby potato.
Keywords: commercialization; native cultivars; native ecotypes; regression; varieties.
Introducción
La papa (Solanum tuberosum L.) es el tercer cultivo alimenticio más importante en el mundo después del arroz y el trigo en términos de consumo humano (Devaux et al., 2014). Se cultiva en más de 100 países, bajo condiciones de climas templados, subtropicales y tropicales, sin embargo, el mejor crecimiento de esta planta se da en regiones frías con noches frescas y días con alta radiación (Mosavi et al., 2018). En los Andes de Sudamérica, el consumo de papa puede alcanzar los 250 kg año-1 per cápita, en estas regiones, la papa constituye el cultivo principal básico, y la mayoría de los hogares cultiva entre 10 a 12 variedades, para reducir su vulnerabilidad a las condiciones ambientales (Alam et al., 2017).
El tubérculo presenta diferentes compuestos bioactivos como; almidón, fibra dietética, aminoácidos, minerales, vitaminas y compuestos fenólicos (Akyol et al., 2016). Los compuestos fenólicos son sintetiza dos por la planta de papa como una respuesta a la protección de bacterias, hongos, virus e insectos (Akyol et al., 2016). Participan en la defensa contra los efectos nocivos de la radiación UV y la protegen de muchos oxidantes (Pervaiz et al., 2017).
La papa con pulpa pigmentada se cultiva entre 2100 msnm hasta 4100 msnm, condiciones en la que resaltan sus características genéticas en la producción de pigmentos naturales en la pulpa del tubérculo, que le otorga un gran contenido de compuestos fenólicos y antioxidantes, y un alto contenido de materia seca (Amorós et al., 2008). Es decir, el tubérculo de pulpa amarilla tiene un elevado contenido de ácido ascórbico (vitamina C) y antioxidantes naturales (Landrum et al., 2001). No obstante, el contenido total de ácido ascórbico no se asocia con el color de la pulpa del tubérculo (Alam et al., 2017). Las variedades de pulpa de color rojo o púrpura contienen antocianinas. La antocianina, es una familia de flavonoides bioactivos hidrosolubles (Liu et al., 2018). Estos compuestos polifenólicos con antocianinas característicos de las papas de pulpa pigmentada tienen propiedades antioxidantes (Kita et al., 2015). Es decir, son ingredientes farmacéuticos potenciales que proporcionan diversos efectos beneficiosos para la salud (Khoo et al., 2017).
A través del mejoramiento se han creado nuevas variedades, con alta capacidad de producción y atributos de mayor calidad para cumplir con los requisitos del mercado, debido a su sabor agradable, digestibilidad y alto valor nutricional, que se consumen en diversas formas (cocidos, horneados, fritos, en forma de sopas, ensaladas, purés) (Soare et al., 2018). Sin, embargo, es escaso su comercialización en forma de minitubérculos o papa baby.
Los minitubérculos generalmente se refiere a su tamaño ya que son más pequeños que los tubérculos semilla convencional, pero más grandes que los tubérculos in vitro (o microtubérculos) producidos en condiciones asépticas (de laboratorio) o en medios artificiales, el tamaño de los minitubérculos puede oscilar entre 5 y 25 mm (Struik, 2007). Los minitubérculos se obtienen generalmente en diferentes métodos (Rykaczewska, 2016). Pero no todos los métodos de obtención de minitubérculos descritos en la literatura pueden ser adecuados para la producción comercial, por lo que los productores de papa deben adaptar las técnicas más actuales; que son las más efectivas para su capacidad (Dimante y Gaile, 2014). Asimismo, aquellos métodos tienen procedimientos costosos (Rykaczewska, 2016). No obstante, la papa se propaga vegetativamente por medio de tubérculos, el material de plantación se llama "tubérculo semilla" y un tubérculo semilla puede producir aproximadamente 10 tubérculos de papa por planta (Kawakami et al., 2015). Siendo un número muy bajo. Además, las políticas de modernización y comercialización de la producción agrícola han puesto en peligro la agrobiodiversidad en los campos de los campesinos (Angé, 2018).
Debido a ello es necesario recurrir por el mejoramiento genético para obtener variedades de pulpa pigmentada que tengan capacidad de producir un alto número de tubérculos por planta. Con alto contenido de compuestos bioactivos y actividad antioxidante (Soare et al., 2018). Debido a que no existen variedades de papa con producción de minitubérculos para el consumo humano, pese a que en el mercado internacional se comercializa como papa coctel, que es la selección de minitubérculos. Es por ello, que este trabajo tuvo como objetivo obtener clones de papa que produzcan minitubérculos con alta calidad nutricional, genética y sanitaria.
2. Materiales y métodos
Localización y condiciones del estudio
El estudio se llevó a cabo en la campiña de Llama, departamento de Cajamarca, ubica- do a 6º22.280’ de Latitud Sur, a 78º51.709’ de Longitud Oeste y a una Altitud de 2900 msnm, con una humedad relativa promedio de 71%, con precipitación anual promedio de 810 mm, el suelo fue franco arcilloso limoso y un pH de 6,3. El ensayo se estableció durante el período lluvioso: campaña (noviembre 2016 - marzo 2017).
Variables estudiadas
Los caracteres biométricos estudiados fueron: (NTT) número total de tubérculos por planta, (NTC) número de tubérculos comerciales por planta, (NTNC) número de tubérculos no comerciales por planta, (RDTO) rendimiento de tubérculos por plan- ta, (PTC) peso de tubérculos comerciales por planta, (PTNC) peso de tubérculos no comerciales por planta, (NT) número de tallos por planta y (AP) altura de planta.
Material vegetal
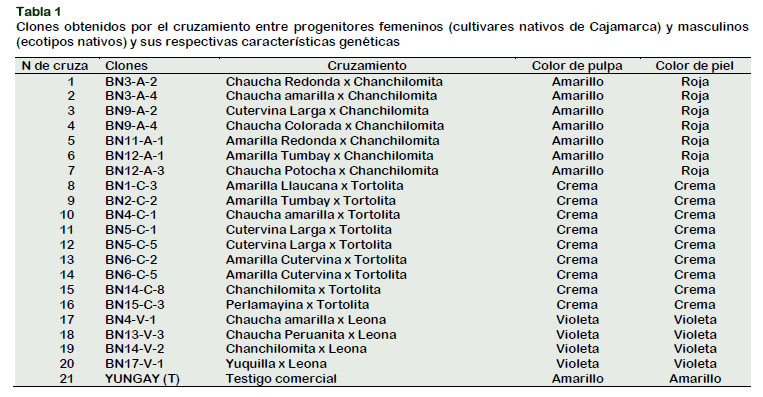
Se evaluaron veinte clones de alta calidad, tolerantes a tizón tardío y pulpa pigmentada, provenientes de las hibridaciones entre cultivares nativos de Cajamarca con tres ecotipos nativos que expresan alto número de minitubérculos. Los progenitores femeninos se seleccionaron de cultivares nativos diploides de pulpa con alto contenido de carotenoides, buena calidad y resistentes a tizón tardío. Por otro lado, la selección de progenitores masculinos se realizó de los ecotipos nativos que producen alto número de tubérculos y su color de pulpa crema, amarilla y violeta; pertenecientes a la especie diploide 2n=2x=24 cromosomas (Solanum phureja Juz et Buck), los cuales se seleccionaron por su bajo contenido de azúcares reductores y su alta producción de tubérculos y la variedad comercial Yungay como testigo (Tabla 1).
Siembra e hibridación de progenitores en invernadero.
Los progenitores fueron cultivados en maceteros dentro de una casa de mallas. En plena floración se colectó el polen de los progenitores masculinos y fue almacenado en recipientes estériles. En los progenitores femeninos se realizaron emasculaciones (eliminación de la antera), posterior- mente se realizaron los cruzamientos correspondientes. Para los cruzamientos se utilizó 20 progenitores femeninos por cada progenitor masculino, cada cruza fue un clon evaluado (Tabla 1).
Cosecha y post cosecha de semilla sexual de papa baby
Las bayas hibridadas por cada clon fueron manejadas adecuadamente hasta la maduración fisiológica, luego cosechadas, maceradas, lavadas, desinfestadas y alma- cenadas en sobres de papel platinado. Se seleccionaron 20 clones en tres grupos, por el color de piel y pulpa (amarillo, crema y violeta) (Tabla 1).
Instalación de semilla sexual en camas de almácigo y evaluación
La semilla sexual debidamente agrupadas, se cultivó en sustratos con alto porcentaje de materia orgánica en camas de almácigo dentro de la casa de mallas. En la cosecha se seleccionaron por el número, peso, color de piel y pulpa de los tubérculos.
Instalación de tubérculos seleccionados en campo experimental y evaluación
Los tubérculos de los 20 clones se instalaron en campo experimental, la unidad experimental fue de 30 plantas (3 surcos de 10 plantas cada uno), siendo considerado el surco central para las evaluaciones. El distanciamiento entre surcos y entre plantas fue de 1 metro x 30 cm, siendo relativamente un menor espaciamiento entre plantas.
Análisis estadístico
El diseño experimental utilizado fue de bloques completos al azar con 21 clones y cada uno con tres repeticiones. Para la comparación entre los promedios se utilizó la prueba de Duncan con un nivel de significación de 0,05%. Para determinar la asociación sobre la característica de importancia (número total de tubérculos por planta) aplicamos el método de regresión múltiple, que permite establecer la mejor relación funcional entre una variable dependiente y las variables independientes para determinar que variable independiente tiene mayor importancia. El análisis estadístico de DBCA y el análisis de regresión múltiple fueron realizados con el paquete estadístico SAS (Statistical Analysis System) (SAS, 2003).
3. Resultados y discusión
Los caracteres biométricos asociados con el número de tubérculos por planta
En la Figura 1, muestra la variación de los clones para el número de tubérculos por planta y sus variables relacionadas a ella.
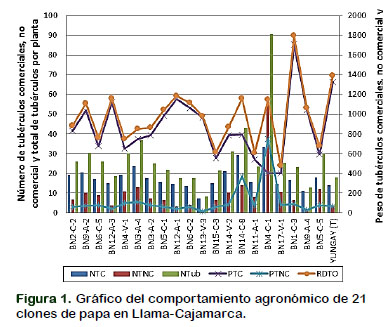
Número total de tubérculos por planta
Los resultados de los cuadrados medios del análisis de varianza de los clones mostrado en la Tabla 2, presentaron diferencias estadísticas altamente significativas para el número total de tubérculos por planta, indicando que los clones tuvieron un comportamiento heterogéneo en el ambiente donde se desarrolló. Estos resultados implican que el medio ambiente influye en el comportamiento diferencial de los clones analizados. Resultados similares concuerdan con lo reportado por Maharana et al. (2017) quienes evaluando la interacción genotipo por ambiente en genotipos de papa, indican la presencia de variación sustancial entre los genotipos sobre el ambiente donde se desarrollan. Mateus et al. (2014), en su investigación realizada en la producción de minitubérculos de papa pone de manifiesto que el ambiente tiene efecto sobre los genotipos y que estos responden de manera distinta en cada ambiente evaluado, demostrando que las condiciones ambientales generadas en cada localidad, son determinantes para la producción de los genotipos.
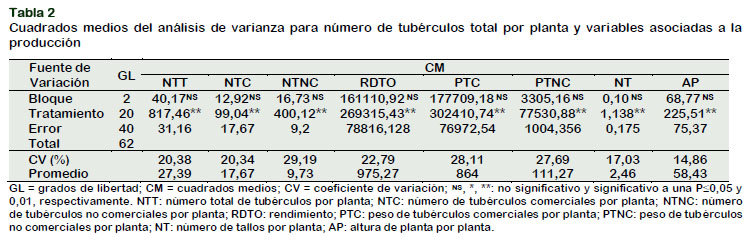
Para la fuente de bloques no presentó significancia indicando que no hubo influencia del medio ambiente dentro del área experimental. El coeficiente de variabilidad (CV), usado como una medida de precisión en la conducción de experimentos fue 20,38%, considerado como aceptable (Tabla 2).
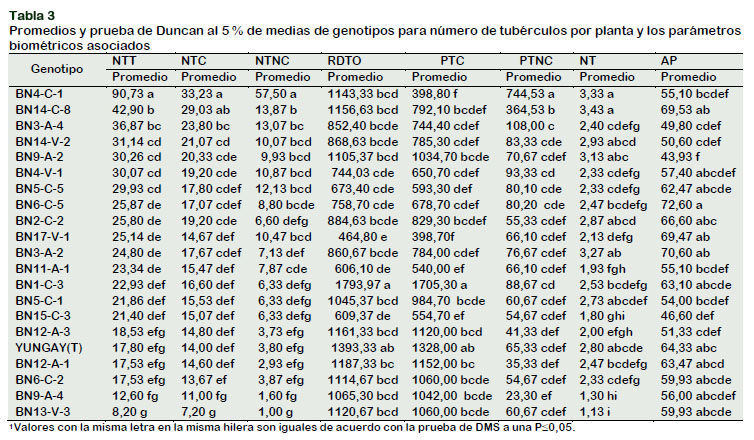
En la Tabla 3, se muestra que el clon BN4-C-1 superó a los demás clones e inclusive a la variedad comercial con 90,73 tubérculos por planta, seguido por BN14-C-8 con 42,9 y BN3-A-4 con 36,87 tubérculos por planta, luego les sigue un grupo homogéneo de doce clones, que va desde el clon BN14-V-2 con 31,14 hasta el clon BN15-C-3 con 21,40 tubérculos por planta, respectivamente. Por último, se presenta un grupo homogéneo de seis clones, quienes reportaron los más bajos promedios siendo el clon BN12-A-3 a BN13-V-3 con promedios que oscilan entre 18,53 a 8,20 tubérculos por planta, respectivamente.
El efecto significativo del aumento de tubérculos por planta fue ligeramente similar a lo encontrado por Rykaczewska (2016), quienes evaluaron cultivares nacionales en aeroponía para la producción de minitubérculos de papa, encontraron que su número de tubérculos por planta fue en promedio entre 32,5 a 36,0 por planta. También resultó similar a los reportado por Tirado (2014), quienes evaluaron la calidad de 18 ecotipos de papa amarilla (Solanum phureja Juz et Buck), encontraron que los genotipos presentan entre 20,43 y 62,24 tubérculos por planta.
Walworth y Carling (2002) indican que el incremento en el número de tubérculos por planta podría afectar su tamaño final. Dagne (2015) reporta que el aumento del número de tubérculos por planta puede deberse al número de tallos por planta producidos a partir de un menor espaciamiento entre plantas, el cual produce un mayor número de tubérculos pequeños. Sin embargo, es un aspecto negativo para producción de tubérculos comerciales, pero positivo para nuestra investigación ya que necesitamos producción de minitubérculos. Por otra parte, el buen tamaño del tubérculo semilla y la alta densidad de plantas produce un alto número de tallos por planta, lo que genera mayor competencia de las plantas por nutrientes, radiación solar entre otros recursos, la cual produce un alto número de tubérculos, sin embargo, los tubérculos producidos serán más pequeños (Shayanowako et al., 2015).
Tubérculos no comerciales por planta
El parámetro biométrico más importante es el número de tubérculos no comerciales por planta debido a la producción de minitubérculos por planta para "papa baby". En este parámetro se recogieron todos los minitubérculos de un tamaño que oscila entre 5 y 25 mm (Struik, 2007).
Los cuadrados medios del análisis de varianza observado en la Tabla 2, detectó diferencias estadísticas muy altas para el número de tubérculos no comerciales por planta. Este resultado indica la presencia de variación sustancial entre los clones sobre los ambientes donde se desarrollan (Maharana, 2017). Para la fuente de bloques no presentó significancia; indicando que no hubo influencia del medio ambiente dentro del área experimental. El coeficiente de variación (CV) fue de 29,19% considerado como aceptable.
La prueba de Duncan al 5% en la Tabla 3, presenta cuatro rangos de significación estadística, compartiendo en el primer rango el clon BN4-C-1 con 57,50 tubérculos no comerciales por planta, la que obtuvo el mayor valor superando estadísticamente a todas los demás clones, seguido por el segundo grupo homogéneo de 7 clones con promedios similares entre sí, que van del BN14-C-8 a BN6-C-5 con valores que oscilan entre 13,87 a 8,80 respectivamente. El tercer rango homogéneo de 10 clones a los que les corresponden el clon BN11-A-1 al clon BN12-A-1 con promedios que varían de 7,87 a 2,93 tubérculos no comerciales por planta y por último las progenies BN9- A-4 con 1,6 y BN13-V-3 con un tubérculo no comercial. Lo que se supone que un alto número tubérculo no comerciales no impli- ca un alto rendimiento. Estos resultados de los clones evaluados fueron consistentes con los resultados obtenidos por Tirado (2014), quienes evaluaron el rendimiento de 32 genotipos avanzados de papa, encontraron que los genotipos presentaron entre 13,61 y 32,53 tubérculos no comerciales por planta.
El número de minitubérculos por planta no solo depende del clon, sino también del número de plantas por área de superficie (Rykaczewska, 2016). A menor espacio entre plantas resulta un mayor número de tubérculos de tamaño inferior a las comerciales (Tesfaye et al., 2013; Abdullah et al., 2016). Del mismo modo Dagne (2015) y Tesfa et al. (2015) señalan que un espaciamiento más estrecho entre plantas, no permite el uso eficiente de nutrientes del suelo y entre otros recursos, el cual se manifiesta en un alto número de minitubér- culos por planta. Sin embargo, el espaciamiento menor entre plantas se debe compensar con distanciamiento más amplio entre surcos (Mahmood, 2005).
El aumento de la densidad de plantas y de tallos por planta, aumenta la cobertura del follaje lo que permite registrar un mayor índice de área foliar y el área fotosintética, lo que favorece la producción y la acumulación de más fotosintatos permitiendo que la planta soporte el desarrollo de un mayor número de tubérculos no comerciales (Abdullah et al., 2016). Estos resultados coinciden con el resultado reportado por Shayanowako et al. (2015), los cuales señalan que las altas densidades de plantas y tallos aumentan el número de tubérculos pequeños. Por lo tanto, estos factores determinan la producción de tubérculos no comerciales los cuales son idóneos para la comercialización como papa baby para el consumo humano.
Tubérculos comerciales por planta
El análisis de varianza para la fuente de clones, fueron altamente significativas (P <0,001) (Tabla 2). Lo cual indica que los clones tuvieron un comportamiento diferencial en el ambiente donde se desarrolló. El coeficiente de variación (CV) fue de 20,34% considerado como aceptable.
La comparación de medias de Duncan (Tabla 3) establece cuatro rangos de significancia estadística, presentando en el primer rango los clones: BN4-C-1 con 33,23 a BN14-C-8 con 29,03, los cuales sobre- salieron con los mayores valores. En el segundo rango se ubicaron los clones: BN3-A-4 a BN6-C-5 con promedios que oscilan entre 23,8 y 17,07 tubérculos comerciales por planta. En el tercer rango presenta el clon BN1-C-3 con 16,6 a BN6-C- 2 con 13,67 y en último rango se encontró a los clones: BN9-A-4 con 11 y BN13-V-3 con 7,2 tubérculos comerciales por planta, respetivamente. Estos resultados coinciden con el resultado reportado por Seminario et al. (2017), quienes evaluando diecisiete cultivares de papa detectaron que el promedio de la planta de papa varió de 9 a 23,57 tubérculos comerciales por planta.
Rendimiento total por planta
En la Tabla 2 se constató que el efecto de los clones para el rendimiento total por planta fue altamente significativo (P < 0,001).
Para la fuente de bloques no pre-sentó significancia en todos los parámetros biométrico evaluados; indicando que no hubo influencia del medio ambiente dentro del área experimental. El coeficiente de variabilidad (CV), usado como una medida de precisión en la conducción de experimentos para el rendimiento fue 22,79%, considerado como aceptable (Tabla 2).
Los resultados de la prueba de Duncan al 5% (Tabla 3), permite confirmar que existen diferencias estadísticas entre los clones, además estableció tres rangos de significación estadística, presentando para el primer rango los clones BNI-C-3 con 1793,97 y YUNGAY (testigo) con 1393,33 g planta-1, los que sobresalieron con mayor rendimiento por planta, para el segundo rango estadístico se encontraron 15 clones: BN12-A-1 al clon BN4-V-1 con promedios de 1187,33 y 744,00 g planta-1. En el último rango se encontró a los clones: BN5-C-5 con 673,4 al BN17-V-1 con 464,8 g planta-1. Resultados similares presentaron por Pérez et al. (2010) en un estudio de interacción de genotipos avanzados, encontraron que el mayor rendimiento de tubérculo varió de 27,47 t ha-1 a 34,46 t ha-1. Seminario et al. (2017) evaluando cultivares de papa reportaron que el rendimiento total de tubérculos por planta varió entre 296 a 987 g planta-1.
Peso de tubérculos comerciales por planta
Los resultados de los cuadrados medios del análisis de varianza de los clones para el peso de tubérculos comerciales por planta mostrado en la Tabla 2, presentaron diferencias estadísticas altamente significativas, indicando que los clones tuvieron un comportamiento heterogéneo en el ambiente donde se desarrolló, para la fuente de bloques no presentó significancia en todos los parámetros biométrico evaluados; indicando que no hubo influencia del medio ambiente dentro del área experimental. Los coeficientes experimentales de variación (CV) fue 28,11%, lo que indica una buena precisión experimental.
Los clones mostraron diferencias estadísticas significativas según Duncan al 5% (Tabla 3), reportando tres rangos estadísticos, donde los valores más altos se manifestaron en los clones BN1-C-3 con 1705,30 y YUNGAY (testigo) con 1328,00 g planta-1 de tubérculos comerciales. Seguido por el rango dos con 14 clones, donde se encuentra los clones: BN12-A-1 con 1152,0 g planta-1 y BN4-V-1 con 650,70 g planta-1 de tubérculos comerciales. En el tercer rango se manifestaron 5 clones: BN5-C-5 a BN17-V-1 con promedios que oscilan entre 593,3 a 398,7 g planta-1 de tubérculos comerciales, respectivamente. Estos resultados coinciden con los reportados por Roa et al. (2010) quienes, evaluando el rendimiento de variedades de papa para el procesamiento industrial, encontraron que el peso comercial de tubérculos por planta varió entre 18,4 y 27,7 t ha-1.
Peso de tubérculos no comerciales por planta
Para el peso de tubérculos no comerciales por planta el análisis de variancia muestra diferencias altamente significativas (P < 0,001), indicando variación entres los clones evaluados. El coeficiente de variación (CV) fue de 27,69% lo que indica una buena precisión experimental.
Los resultados de la prueba de Duncan al 5% (Tabla 3) confirman que entre los promedios de las clones existen diferencias estadísticas, siendo el clon BN4-C-1 la que encabezó el primer rango con un promedio de 744,53 g planta-1 de tubérculos no comerciales, obteniendo mayor peso de tubérculos no comercial, debido a que el clon no alcanzó su madurez comercial, seguido del clon BN14-C-8 con 364,53 g planta-1 de tubérculos no comerciales, siendo superior estadísticamente a los demás clones. En el tercer rango homogéneo se ubicaron los clones: BN3-A-4 a BN15-C-3 con promedios que oscilan entre 108,10 y 54, 67 g planta-1 de tubérculos no comerciales y en el último rango se encontró a los clones BN12-A-3 con 41,33 a BN9-A-4 con 23,30 g planta-1 de tubérculos no comerciales, los que obtuvieron los más bajos valores. Resultados similares fueron descritos por Tirado (2014) quienes evaluaron el rendimiento de 32 genotipos avanzados de papa, encontraron que los genotipos presentan entre 95,32 y 403,81 g planta-1 de tubérculos no comerciales. Dagne (2015) y Mahmood (2005) indicaron que el aumento en la densidad de plantas causa una mayor competencia entre plantas, debido a la disponibilidad de nutrientes para cada planta y como consecuencia resulta en disminución del peso de los tubérculos.
Número de tallos por planta
El análisis de varianza de los cuadrados medios reportó diferencias altamente significativas de los clones para el número de tallos por planta (Tabla 2), indicando que los clones tuvieron un comportamiento diferencias en el ambiente donde se desarrolló, para la fuente de bloques no presentó significancia, indicando que no hubo influencia del medio ambiente dentro del área experimental. Los coeficientes experimentales de variación (CV) fue re- lativamente bajo (17,03%) en el análisis de variancia lo que indica una buena precisión experimental.
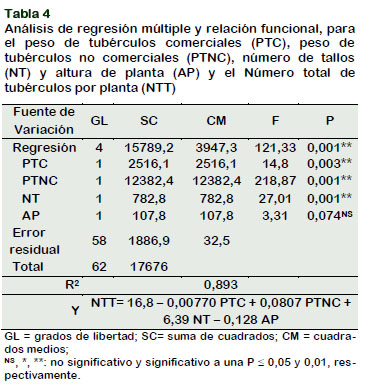
Los clones mostraron diferencias estadísticas significativas en el número de tallos por planta según Duncan al 5% (Tabla 4) esta prueba estableció tres rangos de significación estadística, mostrando con el más alto número de tallos por planta al clon BN14-C-8 con 3,43 compartiendo el primer rango se encontraron también las pro- genies: BN4-23, BN3-A-2, BN9-C-2, BN14-V- 2, BN2-C-2, YUNGAY (testigo) y BN5-C-1 con promedios que oscilan de 3,33 a 2,73. En el segundo rango se encontró a los clones: BN1-C-3 con 2,53 a BN11-A-1 con 1,93. En el último rango los clones BN15-C- 3, BN9-A-4 y BN13-V-3 con 1,8, 1,3 y 1,13 registraron los valores más bajos en el número de tallos por planta, respectivamente. El número de tallos por planta está en función al número de ojos (yemas) del tubérculo, para ello se requiere que el tubérculo semilla tenga mayor resistencia a plagas y enfermedades, tenga buen tamaño, peso, y otros factores; con lo cual se obtendrá un mayor número de tallos por planta y por consiguiente un mejor rendimiento (Tirado, 2014). Núñez (2016) afirman que el número de tallos está ligada a la edad fisiológica de los tubérculos semilla. Asimismo, el número de tallos está relacionado con el tamaño de los tubérculos y la densidad de plantación (Shayanowako et al., 2015). A mayor densidad de tallos, los tubérculos producidos serán más pequeños (minitubérculos) que a bajas densidades de tallos (Shayanowako et al., 2015). Si el tubérculo semilla es demasiado pequeño, tendrá un número bajo de tallos por planta lo que produce sólo unos pocos tubérculos, reduciendo así el rendimiento (Şanlı et al., 2015). Por otra parte, a mayor tamaño del tubérculo semilla producirá un alto número de tallos por planta, que finalmente se traduce en más tubérculos por planta que pueden competir por los recursos del suelo, reduciendo el tamaño del tubérculo, por lo tanto, los mejores tubérculos de siembra son los que son de tamaño medio (Dagne, 2015).
Altura de planta
Los resultados de los cuadrados medios del análisis de varianza de los clones mostrado en la Tabla 2, presentaron diferencias estadísticas altamente significativas para altura de planta, indicando que los clones tuvieron un comportamiento heterogéneo en el ambiente donde se desarrolló, para la fuente de bloques no presentó significancia indicando que no hubo influencia del medio ambiente dentro del área experimental. El coeficiente de variabilidad fue de 14,86%, considerado una buena precisión experimental (Tabla 2).
La prueba de Duncan al 5% para altura de planta se muestra en la Tabla 3. Estableciéndose tres rangos de significación estadística, compartiendo el primer rango doce clones con promedios homogéneos: el clon BN6-C-5 quien presento el mayor valor con 72,6 cm, al clon BN9-A-4 con 56 cm.
Regresión múltiple
La Tabla 4 muestra las características biométricas que más se asocian al número de tubérculos por planta. Se detectaron significancia estadística para el modelo, lo cual indica que existe regresión, sin embargo, no para la variable altura de planta el cual fue no significativo. El coeficiente de regresión indica que el número total de tubérculos (NTT) es influenciado por el peso de tubérculos no comerciales por planta (PTNC), indicando que por cada gramo de peso del tubérculo no comercial se produce un aumento de 0,08 tubérculo por planta, así mismo, está influenciado por el número de tallos (NT), indicando que por cada unidad de tallo por planta se produce un aumento de 6,39 tubérculos por planta. Asimismo, ambas variables presentan una línea de regresión ascendente de izquierda a derecha como se muestra en la Figura 3, lo que implica que conforme se incrementa el peso del tubérculo no comercial y el número de tallos por planta se incrementa también el número total de tubérculos por planta. Por otra parte, el peso de tubérculos comerciales (PTC) y altura de planta (AP), presentaron valores negativos, además se aprecia en la Figura 3, la línea de regresión la misma que es descendente de izquierda a derecha para el PTC y una línea relativamente horizontal para la variable AP, lo que implica que ambas variables no están relacionadas con el NTT.
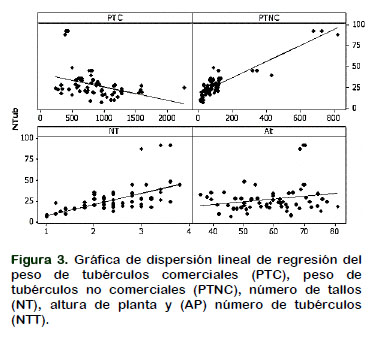
Los resultados de esta investigación concuerdan con los obtenidos por Thornton et al. (2007) quienes evaluando genotipos de papa encontraron que el número de tallos producidos por una planta de papa está directamente relacionado con el número de tubérculos que la planta desarrollará. Debido a que cada tallo tiende a producir cierto número de tubérculos, cuanto más alto sea el número de tallos, más tubérculos serán producidos por cada planta. Por lo tanto, el espaciamiento entre plantas, el tamaño del tubérculo semilla y la densidad de tallos por planta son los factores que determinan la producción de número de tubérculos por planta. A mayor densidad de tallos, los tubérculos producidos serán más pequeños que a bajas densidades de tallo (Shayanowako et al., 2015). Harnet et al. (2014) reportan que a mayor densidad de plantas produce mayor número de tubérculos no comerciales por planta.
4. Conclusiones
El clon que tuvo mayor número de tubérculos por planta fue BN4-C-1 con 90,73 tubérculos por planta y peso promedio de 1143,30 g planta-1, de color de piel crema y pulpa crema, además presentó el mayor número de tubérculos comerciales con 33,23 tubérculos por planta y número de tubérculos no comerciales con 57,50 tubérculos por planta, además del peso no comercial con 398,80 g planta-1, con buen número de tallos por planta con 3,33. Seguido por el clon BN14-C-8 que presentó 42,9 tubérculos por planta y un peso de 1156,63 g planta-1, de piel y pulpa amarilla, así también obtuvo el mayor número de tallos por planta con 3,43. El clon BN3-A-4 presento 36,87 tubérculos/planta y peso de 852,40 g planta-1, 3.0 cm/tubérculo de diámetro, color de piel roja y pulpa amarillo intenso. El clon BN14-V-2 reporto 31,14 tubérculos por planta, 868,63 g planta-1 y color de piel violeta y pulpa amarilla intenso. Siendo estos clones las que obtuvieron una mejor respuesta al mayor número de minitubérculos para la comercialización y consumo humano, como papa baby. El mayor rendimiento por planta fue para el clon BN1-C-3 con 1793,97 g planta-1 y mayor peso comercial con 1705,30 g planta-1. El clon más alto fue para BN6-C-5 con 72,60 cm. La regresión múltiple muestra a la variable número de tallos por planta y peso de tubérculos no comerciales por planta mayor relación funcional con el número de tubérculos por planta.
Se recomienda realizar ensayos de calidad para el procesamiento industrial con los mejores clones resultado de la presente investigación.
Agradecimientos
A la Asociación para la Conservación y Mejoramiento de la Biodiversidad Altoandino - ACOMEBIO, Convenio No. 176-FINCyT-FIDECOM-PIPEI-2012.
Referencias Bibliográficas
Abdullah, M.; Mamun, A.; Mahmud, A.; Zakaria, M.; Mofazzal H.; Hossain, T. 2016. Effects of planting times and plant densities of top-shoot cuttings on multiplication of breeder seed potato. Agriculture and Natural Resources 50: 26-31. [ Links ]
Akyol, H., Riciputi, Y., Capanoglu, E., Caboni, M.F.; Verardo, V. 2016. Phenolic Compounds in the Potato and Its Byproducts: An Overview. Int J Mol Sci. 17(6): 835. [ Links ]
Alam, M.Z., Wang-Pruski, G., Hodges, M., Hawkins, G.R., Kubik, M.D, Fillmore, S.A. E. 2017. Effect of Cooking and Reconstitution Methods on the Loss of Bio- active Compounds in Pigmented and Unpigmented Potatoes. Food and Nutrition Sciences 8: 31-55. [ Links ]
Amorós, W.; Bonierbale, M.; Manrique, K. 2008. Mejoramiento genético de la papa para uso gourmet. Centro Internacional de la Papa CIP. (Lima). [ Links ]
Angé, O. 2018. Interspecies Respect and Potato Con- servation in the Peruvian Cradle of Domestication. Conservat. Soc. 16(1): 30-40. [ Links ]
Dagne, Z. 2015. Influence of seed tuber size and plant spacing on yield and quality of potato (Solanum tuberosum L.) in Holetta, Central Ethiopia. Thesis master of science, Haramaya University, Haramaya. Ethiopia. 106 pp. [ Links ]
Devaux, A.; Kromann, P.; Ortiz, O. 2014 Potatoes for sustainable global food security. Potato Research 57: 185–99. [ Links ]
Dimante, I.; Gaile, Z. 2014. Potato minitubers technology– its development and diversity: A review. Agricultural Sciences (Crop Sciences, Animal Sciences) 1: 69-76. [ Links ]
Harnet, A.; Derbew, B.; Gebremedhin, W. 2014. Effects of inter and intra row spacing on seed tuber yield and yield components of potato (Solanum tubersoum L.) at Ofla Woreda, Northern Ethiopia. Afr.J.Plant SCI. 8(6): 285-290. [ Links ]
Kawakami, T., Oohori, H. and Tajima, K. 2015. Seed potato production system in Japan, starting from foundation seed of potato. Breed Sci. 65(1): 17–25. [ Links ]
Khoo, H.E. Azlan, A., Teng, S.T. and Lim, S.M. 2017. Anthocyanidins and anthocyanins: colored pig- ments as food, pharmaceutical ingredients, and the potential health benefits, Food & Nutrition Research 61: 1-21. [ Links ]
Kita, A., Bąkowska-Barczak, A., Lisińska, G., Hamouz, K., cKuł akowska, K. 2015. Antioxidant activity and quality of red and purple flesh potato chips. LWT - Food Science and Technology 62 (1): 525-531. [ Links ]
Landrum, J.; Bone, R. 2001. Lutein, zeaxanthin and macular pigment. Archives of Biochemistry and Biophysics 385: 28-40. [ Links ]
Liu, L.; Yang, Y.; Gao, J.; Ma, C.; Bi, Y. 2018. A compa- rative transcriptome analysis of a wild purple potato and its red mutant provides insight into the mechanism of anthocyanin transformation. PLoS ONE 13(1): 1-19. [ Links ]
Maharana, J.; Panda, C.; Jakhar, P. 2017. Genotype × Environment Interaction and Stability Analysis of Kharif Potato in Koraput Region of Odisha, India. International Journal of Current Microbiology and Applied Sciences 6(5): 1159-1166. [ Links ]
Mahmood, S. 2005. A Study of Planting Method and Spacing on the Yield of Potato Using TPS. Asian Journal of Plant Sciences, 4: 102-105. [ Links ]
Mateus, R.J.; Haan, S.Z.; Rodríguez, A.D. 2014. Genotype by Environment Effects on Potato Mini- Tuber Seed Production in an Aeroponics System. Agronomy 4: 514-528. [ Links ]
Mosavi, M.; Khorshidi, M.; Masoudian, N.; Hokmabadi, H. 2018. Study of some physiological characteristics of potato tissue under salinity stress. International Journal of Farming and Allied Sciences 7(1): 1-5. [ Links ]
Núñez, P. 2016. Fuentes y dosis de fertilización potásica en el crecimiento, tuberización y calidad de fritura de la papa (Solanum tuberosum L.) variedad única. Tesis de ingeniero, Universidad Nacional Agraria La Molina, Lima. Perú. 110 pp. [ Links ]
Pérez, D.F.; González, A.H.; Mora, O.F.; Rivera, A.P.; Sahagún, J.C.; Balbuena, A.M.; Rubí, M.A.; Gutiérrez, F.R. 2010. Variabilidad genética, diver- sidad fenotípica e identificación de genotipos sobresalientes de papa. [ Links ]
Pervaiz, T.; Songtao, J.; Faghihi, F.; Haider, M.S.; Fang, J. 2017. Naturally Occurring Anthocyanin, Structure, Functions and Biosynthetic Pathway in Fruit Plants. J Plant Biochem Physiol 5(2): 1-9. [ Links ]
Roa, S.; Barboza, C.; Zambrano. A. 2010. Estabilidad del rendimiento de variedades de papa (Solanum tuberosum L.) para procesamiento industrial en el estado Táchira, Venezuela. Revista de la Facultad de Agronomía de la Universidad del Zulia 27: 173- 192. [ Links ]
Rykaczewska, K. 2016. The potato minituber production from microtubersin aeroponic culture. Plant Soil Environ. 62(5): 210–214. [ Links ]
Şanlı, A.; Karadoğan, T.; Erbaș , S.; Tosun, B. 2015. The effects of plant density and eye number per seed piece on potato (Solanum tuberosum L.) tuber yield. Scientific Papers - Series A, Agronomy 58(35): 325- 331.
SAS. 2003. The SAS system for Windows. Release 9.1.3. p. 83-122. SAS Institute, Cary, North Carolina, USA. [ Links ]
Seminario, C.J.; Seminario, C.A.; Domínguez P.A.;Escalante, Z.B. 2017. Rendimiento de cosecha de diecisiete cultivares de papa (Solanum tuberosum L.) del grupo Phureja. Scientia agropecuaria 8(3): 181 – 191. [ Links ]
Shayanowako, A.; Mangani, R.; Mtaita, T.; Upenyu M. 2015. Influence of Main Stem Density on Irish Potato Growth and Yield: A Review. Annual Research & Review in Biology 5(3): 229-237. [ Links ]
Soare, R.; Dinu, M.; Babeanu, C.; Modran, M. 2018. Production of some Colored Potato Genotypes in Organic Culture. Bulletin UASVM Horticulture 75(1): 90-94 [ Links ]
Struik, C. 2007. The canon of potato science: 25. Minitubers. Potato Research 50: 305–308. [ Links ]
Tesfa, B.; Nigussie, D.; Tamado, T. 2015. Influence of Plant Spacing on Seed and Ware Tuber Production of Potato (Solanum tuberosum L.) Cultivars Grown in Eastern Ethiopia. Science, Technology and Arts Research Journal 4(3): 11-17 [ Links ]
Tesfaye, G.; Derbew, B.; Solomon, T. 2013. Combined effects of plant spacing and time of earthing up on tuber quality parameters of potato (Solanum tuberosum L.) at Degem district, North Showa zone of Oromia regional state. Asian Journal of Crop Science 5(1): 24-32. [ Links ]
Thornton, M.; Pavek, M.; Bohl, W.H. 2007. Importance of tuber set and bulking rate. Idaho Potato Conference, India. [ Links ]
Tirado, J. 2005. Determinación de calidad de 18 ecotipos de papa amarilla (Solanum phureja Juz et Buck) en Cutervo-Cajamarca. Tesis de ingeniero, Universidad Nacional Pedro Ruíz Gallo, Lambayeque. Perú. 87 pp. [ Links ]
Tirado, R. 2014. Evaluación del rendimiento de clones avanzados de papa (Solanum tuberosum L.) con pulpa pigmentada – Cajamarca. Tesis de ingeniero, Universidad Nacional Pedro Ruíz Gallo, Lambayeque. Perú. 96 pp. [ Links ]
Walworth, J.L.; Carling, D.E. 2002. Tuber Initiation and Development in Irrigated and Non-Irrigated Potatoes. American Journal of Potato Research 79(6): 387-395. [ Links ]
Received April 4, 2018.
Accepted September 4, 2018.

