










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkScientia Agropecuaria
versión impresa ISSN 2077-9917
Scientia Agropecuaria vol.10 no.1 Trujillo ene./mar. 2019
http://dx.doi.org/10.17268/sci.agropecu.2019.01.12
ARTÍCULOS ORIGINALES
Composición florística de especies herbáceas forrajeras en praderas naturales de las principales microcuencas ganaderas de la región Amazonas
Floristic composition of herbaceous forage species in natural prairies of the main livestock watersheds of the Amazon region
Manuel Oliva* ORCID iD: https://orcid.org/0000-0001-5945-1162; Roicer Collazos; Héctor Vásquez; Karol Rubio; Jorge L. Maicelo
Universidad Nacional Toribio Rodríguez de Mendoza, Chachapoyas, Amazonas, Perú .
Resumen
Existe una gran diversidad de especies forrajeras en las praderas naturales de la región Amazonas, y son escasos los estudios referentes a la identificación de las mismas. Con objeto de profundizar los conocimientos en especies forrajeras, se realizó la identificación de las especies de pastos presentes en las principales microcuencas ganaderas y pisos altitudinales de la región Amazonas. Las microcuencas ganaderas identificadas fueron Shocol, Leyva, Ventilla, Alto Imaza, Pomacochas, Condechaca y Leymebamba. Sobre la base de las microcuencas ganaderas y praderas naturales se definió dos variables referentes a los sistemas de producción (sistemas silvopastoriles , SSP, y sistemas pastoriles a campo abierto , SPCA) y cuatro rangos altitudinales < 1900 m.s.n.m., 1900 – 2400 m.s.n.m., 2400 – 2900 m.s.n.m. y > 2900 m.s.n.m. Se usaron transectos, identificándose un total de 34 familias y 113 especies entre forrajeras y no forrajeras, de las cuales 27 especies fueron clasificadas como especies forrajeras que agrupan un total de 05 familias. Se encontraron diferencias porcentuales de abundancia y diversidad entre especies respecto a las variables estudiadas.
Palabras clave: cobertura; forraje; maleza; Poaceae; zona altoandina.
Abstract
There is a great diversity of forage species in the natural grasslands of the Amazon region, and there are few studies regarding the identification of them. In order to deepen the knowledge on the subject, the identification of the floristic composition of these species in the ma in livestock micro-basins and altitudinal floors of the Amazon region was carried out. The livestock microbasins identified were Shocol, Leyva, Ventilla, Alto Imaza, Pomacochas, Condechaca and Leymebamba. On the basis of livestock micro-basins and natural grasslands, two variables referring to production systems (silvopastoral systems, SSP, and open field pastoral systems, SPCA) and four altitudinal ranges were defined: <1900 m.a.s.l., 1900-2400 m.a.s.l., 2400-2900 m.a.s.l. and> 2900 m.a.s.l. Transects were used, identifying a total of 34 families and 113 species between forages and non -forages, of which 27 species were classified as forage species that group a total of 05 families. We found percentage differences of abundance and diversity among species with respect to the variables studied.
Keywords: cover; fodder; undergrowth; Poaceae; high Andean zone.
1. Introducción
Según el IV Censo Nacional Agropecuario (INEI 2012), la región Amazonas cuenta con 157,166 cabezas de ganado vacuno, convirtiendo la actividad ganadera en una de las actividades de mayor importancia económica, la misma que en los últimos años ha tenido un notorio incremento, principalmente en las provincias de la zona sur como son Chachapoyas, Bongará, Rodríguez de Mendoza y Luya.
La alimentación del ganado en la zona se sustenta básicamente en el cultivo de pastos y manejo de praderas naturales, estas últimas representan más del 70% del área destinada al pastoreo las cuales se han ido formando sobre áreas de bosques primarios, abiertas para instalar cultivos de pan llevar bajo el sistema de "agricultura migratoria", en las cuales por efectos naturales proliferan especies herbáceas nativas con aptitud forrajera, cuya composición y clasificación taxonómica son desconocidas y ha sido el objeto del presente estudio (IIAP et al., 2007).
El N2O y el CH4 emitido por los rumiantes son los GEI más importantes y no solo se relaciona con problemas ambientales, amenaza la sustentabilidad de los sistemas de producción debido a una disminución en su eficiencia productiva (Moumen et al., 2016), pero teniendo en cuenta que el uso de forrajes de mejor calidad genera beneficios nutricionales y ambientales ya que se reducen menores cantidades de metano (CH4) y se mejora la nutrición animal, debido a la mejor conformación de los carbohidratos de los forrajes (Melesse et al., 2017; Patra, 2016), es de resaltar el importante aporte de las praderas a la conservación del medio ambiente, ya que se constituyen en sumideros de carbono debido a sus sistemas radiculares y si capacidad fotosintética (Berninger et al., 2015).
En ese sentido, gracias a la biodiversidad existente en la zona de estudio, es posible utilizar especies nativas con aptitud forrajera que demuestren ventajas comparativas y a su vez sirvan para los programas de mejoramiento genético que permitan aprovechar sus características de adaptación, características agronómicas como el ciclo productivo, rendimiento estable, resistencia a enfermedades y otros aspectos, cuyos estudios tienen como primer punto la identificación y distribución de las especies que en la actualidad tiene como nicho ecológico a las praderas naturales ubicadas en las distintas microcuencas ganaderas de la región Amazonas, bajo distintos sistemas de producción ganadera.
El objetivo de esta investigación es determinar la composición florística de especies herbáceas forrajeras de las principales cuencas ganaderas de la región Amazonas teniendo en cuenta dos sistemas de producción: sistemas silvopastoriles y sistemas a campo abierto en 4 pisos altitudinales.
2. Materiales y métodos
Las áreas de estudio de praderas naturales, se encuentran en el ámbito de la región Amazonas, en las cuencas ganaderas de Shocol, Leyva, Ventilla, Alto Imaza, Pomacochas, Condechaca y Leymebamba, presentan una temperatura media anual de 16°C, con temperatura mínima de 10 y máxima de 20°C. Las zonas identificadas comparten condiciones ambientales, textora de suelo, estructura y drenaje similares. Las especies herbáceas fueron muestreadas en el periodo mayo septiembre del 2015. Para clasificar las especies forrajeras y no forrajeras se recurrió a la experiencia de los productores y personas del lugar, así como de técnicos de campo. En la etapa de identificación y delimitación de microcuencas ganaderas, la información base que se utilizó fue imágenes satelitales ETM+ con una resolución de 30 metros, obtenidas del satélite LANDSAT, La cartografía utilizada fue dos cartas nacionales la 12H y 13H del IGN (Instituto Geográfico Nacional) en coordenadas UTM, con datum WGS 84, en la zona 18, se obtuvo datos de ríos, quebradas, curvas a nivel, información que fue superpuesta sobre las imágenes satelitales. El trabajo de digitalización se realizó a partir de la delimitación de las microcuencas ganaderas, en base a la información cartográfica del IGN y las imágenes satelitales, la demarcación de las zonas se realizó con el software ArcGis v.10.1.
Para realizar el muestreo de las especies herbáceas, se utilizó el método del transecto lineal, el cual consiste en estirar una cuerda delgada sobre la pradera con 100 marcas cada 50 cm denominados puntos de contacto, tratando que la cuerda corte todas las características posibles de la pradera (espacios de alta, baja y mediana densidad, así como suelos pobres, fértiles y demás características que pueda tener la pradera). Se eligió al azar un punto de inicio para luego registrar las especies en cada punto de contacto. Como regla general se determinó una repetición por cada hectárea de pradera natural identificada, tanto para el caso del sistema silvopastoril como para el sistema pastoril a campo abierto. Con la ayuda de una cartilla de registro y en función a los puntos de contacto se toma los datos según sea el caso la información correspondiente a: nombre común, su clasificación como grupos fisonómicos, funcionales por ejemplo "gramíneas", "leguminosas" y "otras especies" o de acuerdo a su aptitud como "forrajera" o "no forrajera", esta última corresponden a la proyección de malezas presentes en la pradera. Las especies desconocidas fueron cuidadosamente recolectadas y deshidratadas haciendo uso de prensas botánicas y periódicos en una estufa a 48 °C durante 168 horas; posteriormente las muestras fueron enviadas a un laboratorio especializado para su caracterización botánica de especies herbáceas. La clasificación botánica se realizó en el Herbario Nacional de Trujillo (HUT), entidad que pertenece a la Universidad Nacional de Trujillo y es de amplia experiencia y reconocida trayectoria en botánica del Perú.
3. Resultados y discusión
Se identificaron 34 familias y 113 especies, de las cuales 27 especies fueron clasificadas como especies forrajeras que se agrupan en cinco familias (Tabla 1).

De las especies forrajeras identificadas la familia mejor representada fue la Poaceae con 22 especies distintas, seguida de la familia Asteraceae con dos especies, además de las familias Fabaceae, Caryophyllaceae y Commelinaceae con una especie por cada una de ellas. En un estudio similar en las cuencas de Molinopampa, Chachapoyas y Leymebamba Oliva et al. (2015) identificaron 11 especies del conocimiento común de los lugareños, utilizadas como pastos, ubicándolos en cinco familias, Asteraceae, Poaceae, Fabaceae, Polygonaceae a Commelinaceae.
A pesar de la diversidad de especies encontradas cabe destacar que en su gran mayoría se trata de malezas, lo que equivale a decir pastos de mala calidad para el ganado, sin embargo, esta situación es reversible realizando prácticas de manejo que logren compatibilizar la conservación y producción de las praderas naturales, manejando la pradera con intensidades de pastoreo intermedias, logrando de esta manera optimizar tanto la producción animal como vegetal (Justo, 2015).
Así mismo, considerando la variable altitud, en el rango altitudinal de 1400 – 3400 m.s. n.m. se encontraron las especies forrajeras (Tabla 2).
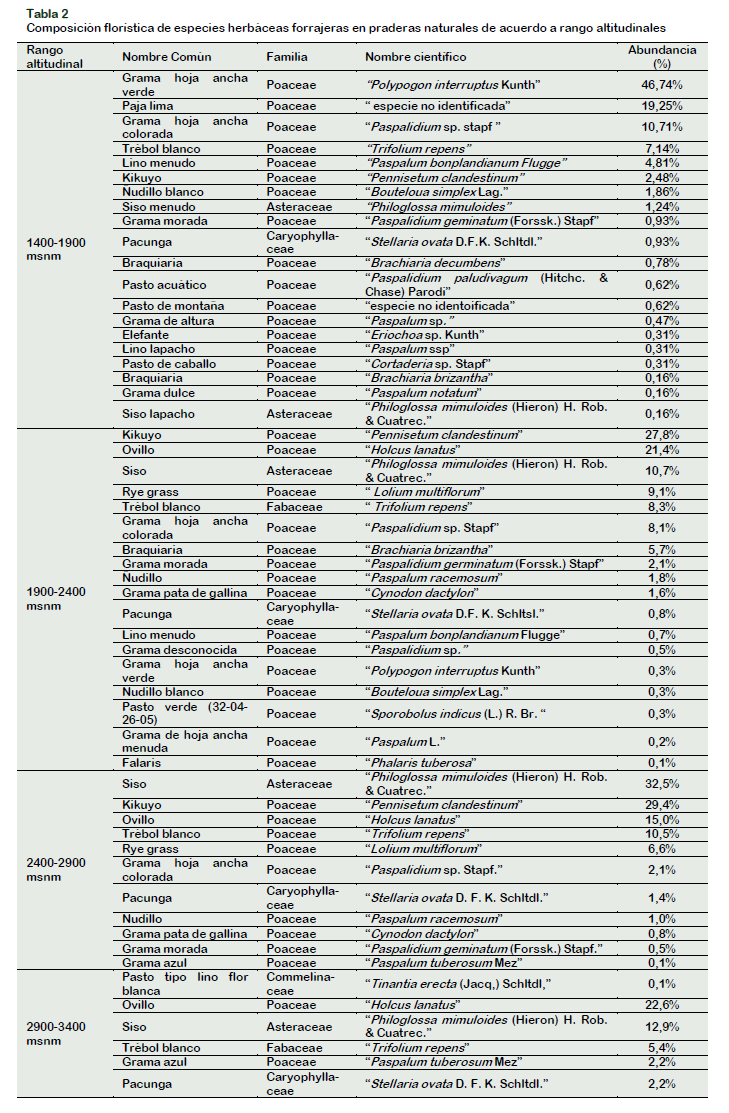
En referencia a los rangos altitudinales, los resultados encontrados fueron muy variados: en el primer rango altitudinal la especie Polypogon interruptus Kunth "Grama de hoja ancha verde" es la más abundante con un 46%; en el segundo rango altitudinal la especie Pennisetum clandestinum "Kikuyo" fue la más abundante con un 27,8%; en el tercer rango altitudinal la especie más abundante es Philoglossa mimuloides (Hieron.) H. Rob. & Cuatrec. "Siso" con un 32,5% y en el cuarto rango altitudinal que corresponde a las zonas de mayor altitud la especie más abundante fue Jarava ichu Ruiz & Pav. "Ichu".
Además, se identificaron dos especies que fueron Trifolium repens "Trébol blanco" y Philoglossa mimuloides (Hieron.) H. Rob. & Cuatrec. "Siso" las cuales se encontraron en todos los rangos altitudinales, con mayor abundancia en los pisos altitudinales más altos. Un resultado atractivo respecto a la variable altitud es la capacidad de algunas especies como Pennisetum clandestinum "Kikuyo" y Philoglossa mimuloides "Siso" para adaptarse a diferentes pisos altitudinales. La primera de estas fue identificada en un punto más bajo a 1585 m.s.n.m. en el distrito de Mariscal Benavides, microcuenca ganadera Leyva, y en un punto más alto a los 2877 m.s.n.m. en el distrito de Olleros, microcuenca ganadera del Alto Imaza, mientras que la segunda especie se encontró en su punto más bajo a los 1633 m.s.n.m. en el sector denominado Challuayacu, distrito de Limabamba, microcuenca ganadera Shocol, y su punto más alto a los 3175 m.s. n.m. en el sector denominado Ipaña, distrito de Leymebamba, microcuenca ganadera de Leymebamba.
En el rango altitudinal más bajo (1400-1900) se encontró mayor diversidad florística respecto a otros estratos más altos, esto coincide con lo reportado por Ramírez (2013), quien expresa que a medida que se incrementa la altura disminuye el número de especies encontradas. Por otra parte, en el rango altitudinal más alto que fue de 2900 m s. n. m. en adelante la especie dominante fue Jarava ichu Ruiz & Pav. perteneciente a la familia de las Poaceae, esta especie está presente en muchas de las zonas altoandinas del país, según lo corroboran estudios como los de Argote et al. (2013), además reportó que la familia con mayor valor de importancia para el rango de 3600-3800 msnm fue la famila Poaceae y a rangos altitudinales más altos tenían dominio otras familias como Asteraceae, resultados coincidentes con los datos obtenidos en este estudio, ya que la familia dominante también fue la Poaceae hasta los 3400 m s. n. m. aproximadamente. Si bien es cierto, no se estudiaron cuencas ganaderas en pisos altitudinales más altos, pero se evidenció que a medida que iba aumentando la altura disminuía la diversidad de especies de esta familia en particular.
Un dato curioso es que la especie Trifolium repens "Trébol blanco" perteneciente a la familia de las Fabaceaes es una especie con gran adaptación, ya que se encuentra presente en todos los pisos altitudinales, con mayor abundancia en los pisos altitudinales más altos lo que difiere con lo reportado por Cabrera y Rivera (2016) quienes obtuvieron valores altos del índice de valor de importancia para las Fabaceaes en un rango altitudinal bajo que estuvo entre 127 y 187 m s. n. m. y con lo obtenido por Mora et al. (2013) quienes encontraron mayor diversidad de esta familia a una altitud de 366 m s. n. m., lo que se explicaría por la mayor duración que presenta su estación lluviosa en el primer caso y por el largo periodo de barbecho en el segundo caso; ambos factores determinantes para la existencia de una mayor diversidad florística. Sin embargo, autores como Valenzuela et al. (2015) reportaron esta familia en pisos altitudinales por encima de los 1400 m s. n. m.
De igual forma se obtuvo la composición florística de especies herbáceas forrajeras teniendo en cuenta la variable microcuenca ganadera (Tabla 3).
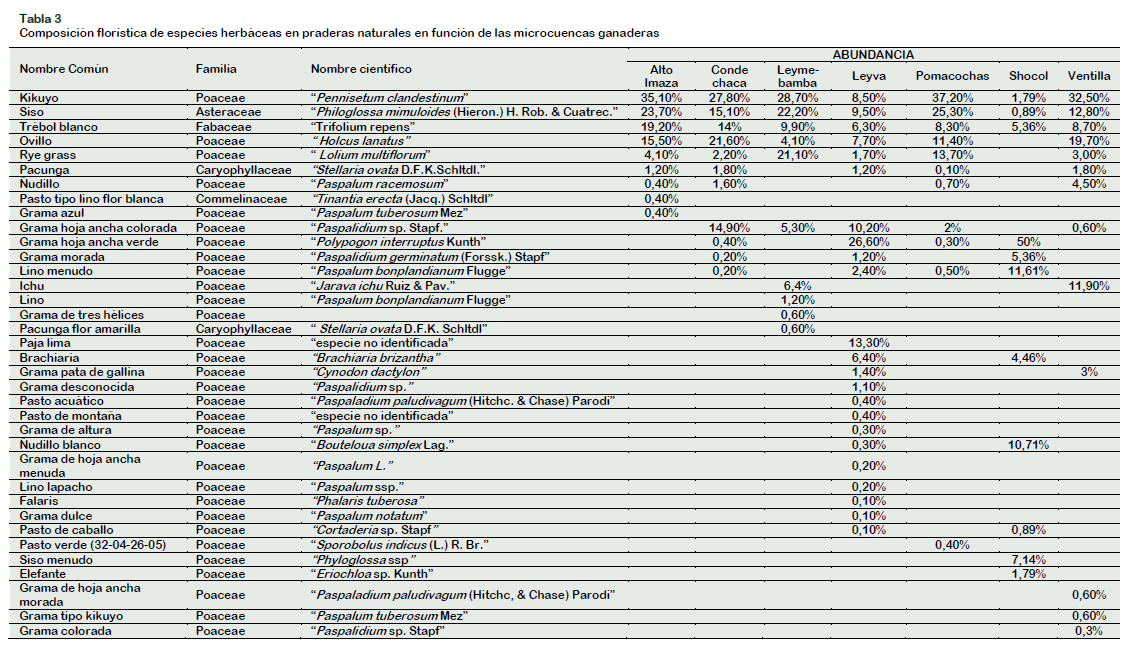
De acuerdo a esta variable, la especie forrajera Pennisetum clandestinum comúnmente llamada "Kikuyo" es la especie más abundante en las microcuencas ganaderas Alto Imaza, Condechaca, Leymebamba, Pomacochas y Ventilla con porcentajes mayores a 27%; mientras que en las microcuencas ganaderas Shocol y Leyva la especie forrajeras Polypogon interruptus Kunth "Grama de hoja ancha verde" es la especie más abundante con 50% y 26,6% respectivamente, notándose que dicha especie es más abundante en zonas tropicales las cuales abarcan gran parte del área destinada a la ganadería en las microcuencas ganaderas en mención.
Las especies forrajeras Pennisetum clandestinum "Kikuyo", Philoglossa mimuloides "Siso" y Trifolium repens "Trébol blanco", pertenecientes a las familias botánicas Poaceae, Asteraceae y Fabaceae respectivamente se encontraron en todas las microcuencas estudiadas, con porcentajes de abundancia de 24,5%; 15,7% y 10,3%, notándose claramente su adaptación a diferentes pisos altitudinales, climas y tipos de suelo; también destaca con las mismas cualidades que las especies citadas, la especie forrajera Holcus lanatus "Ovillo" la cual se encontró en seis de las siete microcuencas estudiadas.
Como se aprecia la familia de especies más representada dentro de las siete microcuencas es la familia Poaceae, las cuales no sólo destacan por su abundancia dentro de las praderas sino también por la diversidad de especies existentes. Sin embargo, en términos de abundancia destaca en un segundo lugar la especie Philoglossa mimuloides (Hieron.) H. Rob. & Cuatrec. "Siso" de la familia botánica Asteraceae, con un porcentaje promedio de 15,7% dentro de las siete microcuencas ganaderas estudiadas. En las microcuencas ganaderas de Shocol y Leyva la especie forrajera más abundante es Polypogon interruptus Kunth, notándose de esta manera la preferencia de la especie por zonas tropicales, las cuales abarcan gran parte del área destinada a la ganadería en las microcuencas ganaderas en mención. Cabe destacar además que la microcuenca ganadera Leyva, fue la que presentó mayor diversidad de especies forrajeras, esto se debe principalmente a que involucra muchos pisos altitudinales que van desde los 1600 m s. n. m. en el distrito de San Nicolás, hasta los 2800 m s. n. m. en la naciente del río San Antonio en el sector denominado Ocol, distrito de Molinopampa.
Por otro lado, teniendo en cuenta la variable sistema de producción: SSP o SPCA, los resultados encontrados fueron los siguientes: (Tabla 4).
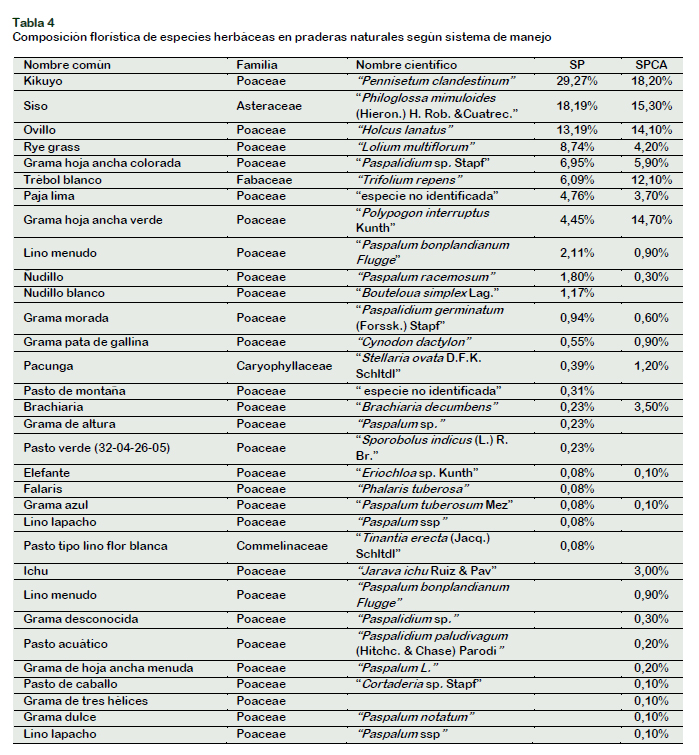
Como se puede observar la composición florística de especies herbáceas forrajeras bajo SSP incluye un total de 23 especies distribuidas en cinco familias botánicas que son Poaceae, Asteraceae, Fabaceae, Caryoghyllaceae y Commelinaceae, de las cuales las Poaceas y las Asteraceaes son las más representativas con las especies Pennisetum clandestinum y Philoglosa mimuloides, con porcentajes de 29,27% y 18,19%, ocupando el primer y segundo lugar respectivamente. La familia dominante para ambos sistemas de explotación ganadera tanto sistemas silvopastoriles como sistemas pastoriles a campo abierto fue la Poaceae, datos coincidentes con los obtenidos por Gandullo et al. (2013) y Ramírez (2013).
Por su parte bajo SPCA se identificaron cuatro familias botánicas: Poaceae, Asteraceae, Fabaceae y Caryophyllaceae. De ellas la más representativa fue la familia Poaceae, cuya especie más abundante fue Pennisetum clandestinum "Kikuyo" con 18,2%, en segundo lugar, encontramos a la especie Philoglosa mimuloides "Siso" perteneciente a la familia Asteraceae con un 15,3% de abundancia. Las especies Polypogon interruptus Kunth "Grama hoja ancha verde" y Holcus lanatus "Pasto ovillo" pertenecientes a la familia Poaceae, ocupan el tercer y cuarto lugar en términos de abundancia con 14,7% y 14,1% respectivamente y en el quinto lugar encontramos a la especie Trifolium repens "Trébol blanco" perteneciente a la familia Fabaceae, con un 12,1% de abundancia.
El porcentaje de abundancia de determinada especie también está en función a la carga animal y al tipo de sistema de explotación ganadera en el que se encuentren, por ejemplo, bajo un sistema a campo abierto la diversidad de familias es mucho menor que en un sistema silvopastoril (Sánchez et al., 2005). Es importante tener en cuenta que a pesar de que cada tipo de sistema de explotación ganadera, ya sea sistema silvopastoril, sistema a campo abierto, sistema estabulado, sistema semiestabulado tiene sus ventajas y desventajas, es evidente que la actividad pecuaria va a ejercer un impacto en los ecosistemas y éste resulta negativo si la realización de estas actividades compromete la sustentabilidad de los recursos naturales presentes (Molina et al., 2013).
Existe un factor muy importante que no podemos dejar de lado cuando se trata de composición florística, el factor ambiental, ya que según indican Gandullo et al. (2013), las condiciones ambientales junto con el tipo de manejo de los recursos naturales de un determinado lugar son los que determinan la producción primaria neta y la abundancia de las especies que lo constituyen.
Otro factor a considerar es la época en que se realicen los muestreos; ya que si se hace en época seca como fue en este caso la calidad y abundancia de los pastos tanto nativos como naturales va a ser menor (Villalta et al., 2016).
4. Conclusiones
Existe una gran diversidad florística en las praderas naturales de Amazonas, sin embargo, son relativamente pocas las especies que se consideran forrajes y además son considerados aun malezas, por lo que son un forraje de limitada calidad para el ganado.
Es de notar que esta diversidad se reduce en función a la altura en la que se encuentran las praderas naturales, así, a mayor altura menor es la diversidad y es diferente así mismo la especie más representativa, por ejemplo, a menor altura es más representativa la especie Pennisetum clandestinum, mientras que, a mayor altura, por encima de los 2800 msnm Jarava ichu es la más representativa.
Se comprueba la existencia de especies importantes como "siso", "kikuyo" y "trébol blanco" que a pesar de tener relevancia en las praderas ganaderas no son de alto valor nutritivo, lo que lleva a recomendar mayor intervención en el mejoramiento de estos pastos.
Referencias bibliográficas
Argote, G.; Aguirre, L.; Flores, E. 2013. Frecuencia de Trifolium amabile Kunth (Fabaceae) en dos sitios del altiplano de Puno, Perú. Ecología Aplicada 12 (2): 83-89. [ Links ]
Berninger, F.; Susilouto, S.; Gianelle, D.; Bahn, M.; Wohlfahrt, G.; Sutton, M.; Garcia-Pausas, J.; Gimeno, C.; Sanz, M.J.; Dore, S.; Rogiers, N.; Furger, M.; Eugster, W.; Balzarolo, M.; Sebastià, M.T.; Tenhunen, J.; Staszewski, T.; Cernusca, A. 2015. Management and site effects on carbon balances of European mountain meadows and rangelands. Boreal Environment Research 20: 748-760. [ Links ]
Cabrera, D.M.; Rivera, O. 2016. Composición florística y estructura de los bosques ribereños de la cuenca baja del río Pauto, Casanare, Colombia. Caldasia 38(1): 53-85. [ Links ]
Gandullo, R.; Fernández, C.; Schmid, P.; Giménez, G. 2013. Efecto de la variación ambiental en la flora, vegetación y productividad de los humedales "Mallines" de la provincia de Neuquén. Argentina. Revista de la facultad de agronomía UNL Pam 22(2): 75-83. [ Links ]
IIAP (Instituto de Investigaciones de la Amazonía Peruana); GRA (Gobierno Regional Amazonas); ECOAN (Asociación Ecosistemas Andinos). 2007. Propuesta de zonificación ecológica y económica del departamento de Amazonas. IIAP, Amazonas, Perú [ Links ].
INEI (Instituto Nacional de Estadística e Informática). 2012. IV Censo Nacional Agropecuario. Sistema de consulta de datos. [ Links ]
Justo, C. 2015. Composición florística y especies indicadoras de la familia Asteraceae en la región de los campos del noreste de Uruguay y sur de Brasil. Tesis de licenciatura, Universidad de la República, Uruguay. 30 pp. [ Links ]
Melesse, A.; Steingass, H.; Schollenberger, M.; Holstein, J.; Rodehurtscord, M. 2017. Nutrient compositions and in vitro methane production profiles of leaves and whole pods of twelve tropical multipurpose tree species cultivated in Ethiopia. Agroforestry Systems 91: 1-13. [ Links ]
Molina, V.M.; Pando, M.; Alanís, E.; Canizales, P.A.; González, H.; Jiménez, J. 2013. Composición y diversidad vegetal de dos sistemas de pastoreo en el matorral espinoso tamaulipeco del Noreste de México. Revista Mexicana de Ciencias Pecuarias 4(2): 361-371. [ Links ]
Mora, C.A.; Alanís, E.; Jiménez, J.; González, M.A.; Yerena, J.I.; Cuellar, L.G. 2013. Estructura, composición florística y diversidad del matorral espinoso Tamaulipeco, México. Ecol. Apl. 12(1): 29-34. [ Links ]
Moumen, A.; Azizi, G.; Chekroun, K.B.; Baghour, M. 2016. The effects of livestock methane emission on the global warming: Revisión. International Journal of Global Warming 9: 229-253 [ Links ]
Oliva, M.; Oliva, C.; Rojas, D.; Oliva, M.; Morales, A. 2015. Identificación botánica de especies nativas de pastos más importantes de las cuencas lecheras de Molinopampa, Pomacochas y Leymebamba, Amazonas, Perú. Scientia Agropecuaria 6(2): 125–129. [ Links ]
Patra, A.K. 2016. Recent advances in measurement and dietary mitigation of enteric methane emissions in ruminants. Frontiers in Veterinary Science: 3-39 [ Links ]
Ramírez, M.G. 2013. Diversidad florística a diferente altitud en el ecosistema páramo en siete comunidades de la OSG UNOCANT. Tesis para obtener el grado de ingeniero forestal, Escuela Superior Politécnica de Chimborazo-Ecuador. 125 pp. [ Links ]
Sánchez, D.; Harvey, C.A.; Grijalva, A.; Medina, A.; Vílchez, S.; Hernández, B. 2005. Diversidad, composición y estructura de la vegetación en un agropaisaje ganadero en Matiguás, Nicaragua. Rev. Biol. Trop. 53(3): 387-414. [ Links ]
Valenzuela, L.M.; Ríos, J.C.; Barrientos, K.R.; Muro, G.; Sánchez, J.; Briceño, E.A. 2015. Estructura y composición florística en dos comunidades de Mezquite (Prosopis laevigata (Humb. & Bonpl. Ex willd.) M.C. Johnst) en Durango, México. Interciencia 40(5): 465-472. [ Links ]
Villalta, P.I.; Zapana, J.G.; Zapana, J.C.; Araoz, J.; Escobar, F. 2016. Evaluación de pastos y capacidad de carga animal en el fundo "Carolina" de la Universidad Nacional del Altiplano-Puno Perú. Rev. Investig. Altoandin. 18(2): 303-310. [ Links ]
Received June 21, 2018.
Accepted March 2, 2019.
* Corresponding author
E-mail: manuel.oliva@untrm.edu.pe (M. Oliva).

