










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkScientia Agropecuaria
versión impresa ISSN 2077-9917
Scientia Agropecuaria vol.10 no.2 Trujillo abr./jun. 2019
http://dx.doi.org/10.17268/sci.agropecu.2019.02.03
ARTÍCULOS ORIGINALES
Inhibitory action of dried leaf of Cassia alata (Linn.) Roxb against lipoxygenase activity and nitric oxide generation
Babatunde Joseph Oso1,* ORCID iD https://orcid.org/0000-0002-5394-430X; Kayode Olayele Karigidi2 ORCID iD https://orcid.org/0000-0001-8667-7108
1 Department of Biochemistry, McPherson University, Seriki-Sotayo, P.M.B 2094, Abeokuta, Ogun State, Nigeria.
2 Department of Biochemistry, Ondo State University of Science and Technology, P.M.B 353, Okitipupa, Ondo State, Nigeria.
Abstract
The mechanisms of inflammation mediated by metabolites of lipoxygenase and nitric oxide play essential roles in physiological immune response. The aim of this study was to evaluate the lipoxygenase (LOX) inhibitory activity of methanolic extract of dried leaf of Cassia alata (Linn.) Roxb. as well as its effect on in vitro accumulation of nitric oxide (NO). The LOX inhibitory activity was evaluated for its ability to inhibit lipoxygenase (soyLOX) by absorbance-based assay using linoleic acid as the substrate while the influence of the extract on accumulation of NO was monitored spectrophotometrically by Griess assay method. Moreover, gas chromatographic analysis was carried out on the ether extract to identify and quantify the volatile compounds in the extract. The study showed effective soyLOX inhibitory activity correlating with the NO depletion in a concentration- dependent manner. The gas chromatographic identification of the volatile compounds in the extract showed the presence of stearic, oleic and linoleic acids. This study revealed the methanolic extract of the leaf of C. alata (Linn.) Roxb. as potent inhibitor of both soyLOX and nitric oxide generation. Thus, these results give indications regarding therapeutic interest of C. alata (Linn.) Robx. as a potential anti- inflammatory agent.
Keywords: Cassia alata (Linn.) Roxb.; lipoxygenase; nitric oxide; inflammation; gas chromatography.
1. Introduction
Inflammation occurs as cellular and molecular reactions which play vital roles against pathogenic agents and tissue damage (Medzhitov, 2008). The mecha- nisms of inflammatory responses involve series of events in which the metabolism of arachidonic acid plays a central role by the concerted and regulated actions of cyclo- oxygenase (COX) and the lipoxygenase (LOX) in the syntheses of biologically active mediators such as prostaglandins, throm- boxane A2, hydroperoxyeicosatertraenoic acids (HPETE’S) and leukotrienes (LT’s) (Alitonou et al., 2006; Rådmark et al., 2015). These eicosanoids have varied biological functions which include the enhancement of inflammatory response through promotion of oedema formation and recruitment of leukocytes to the sites of inflammation (Serhan, 2002; Dennis and Norris, 2015). Moreover, improper regula- tion of these metabolites had been associated with the development of various diseases such as cancer, arthritis, chronic bronchitis, atherosclerosis and asthma (Li et al., 2013; Wisastra and Dekker, 2014). Inhibition of COX and LOX had been shown to ameliorate disease progression in different experimental models (Sarah et al., 2015). However, other mediators of inflammatory responses could interfere with the metabolites of LOX and the respective signalling pathways thereby sustaining or abating the cascade of events leading to their respective biological effects. One of such mediators is nitric oxide (NO) whose participation had been found to potentiate a number of pathologies that are associated with HPETE (Canals et al., 2003). In addition, NO is known as a biomarker of airway inflammation greatly associated with cysteinyl-leukotrienes, the products of LOX (Larsson et al., 2005). Moreover, inhibition of NO generation had been shown to result in anti-inflammatory activity in macrophages (Khlifi et al., 2013). Hence, inflammatory disorders are complex diseases whose treatment should involve more than one target and thus development of polyfunctional drugs.
Anti-inflammatory drugs targeting inhibition of LOX and NOS have been developed for therapeutic indications. However, extensive and prolonged use of these therapeutics is usually coupled with wide range of adverse effects which had led to the exploration of botanicals with such properties in the prevention and treatment of inflammatory disorders. In the quest to search for medicinal plants with such multifunctional properties, the present study was planned to evaluate the LOX inhibitory and NO scavenging properties of the methanolic extract of the dried leaf of Cassia alata (Linn.) Roxb., a shrub belonging to the family Leguminosae- Caesalpiniacea with therapeutic potentials commonly cultivated as an ornamental plant and used in folk medicine for the treatment of various pathologies such as skin infection and scabies (Oso and Olowookere, 2018). The botanical has been reported to contain wide range of compounds such as phytosterols, fatty acids, terpenes, esters and alcohols with potent antioxidant activities (Chua et al., 2019).
The objective of this study was to inves- tigate the inhibitory action of dried leaf of Cassia alata (Linn.) Roxb against lipoxy- genase activity and nitric oxide generation
2. Materials and methods
Preparation of plant sample
Dried leaf of C. alata (Linn.) Roxb. (50 g) was soaked in methanol (200 mL) for 24 hours, after which the sample was filtered and dried. The dried sample was then reconstituted in methanol to a concen- tration of 1 mg/mL, making a stock solution from which appropriate dilutions were prepared for subsequent analyses.
Soybean lipoxygenase (soyLOX) inhibitory activity
Crude lipoxygenase from soybean was prepared as reported by Leelaprakash et al. (2012). Pulverised soybean was defatted and decolorized with cold acetone. The defatted sample was dried overnight. Exactly 10 g of the pulverised sample was added into 30 mL of 50 mM sodium phosphate buffer (pH 6.8) and constantly stirred for 5 hours at 40 °C to form a slurry. The slurry was filtered through mesh cloth. The filtrate was centrifuged at 12000 rpm for 15 minutes using a refrigerated centri- fuge and the supernatant was subsequently used as LOX source. The anti-lipoxygenase activity was studied using linoleic acid as the substrate and soyLOX as the enzyme according to the method described by Shinde et al. (1999). Exactly 250 µL of the sample at various concentrations of 0, 25, 50, 75 and 100 µg/mL was mixed with 250 µL of 0.1 M phosphate buffer (pH 9.0) and 150 µL of soyLOX. The mixture was allowed to stand at room temperature of 30 °C for 5 minutes. Then, 500 µL of 0.6 mM linoleic acid solution was added and mixed gently. The absorbance was read at 234 nm. Indo- methacin was used as reference standard. The percent inhibition was calculated using the following equation:
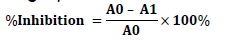
Nitric oxide Accumulation Assay
Nitric oxide radical scavenging activity was determined according to the method reported by Garrat (1964). The principle is based on the reaction between nitrite (generated spontaneously from sodium nitroprusside in aqueous solution at physiological pH) with sulfanilic acid at low pH to form diazonium ion, which couples with α-naphthylamine producing a red violet azo dye measurable at 540 nm. Exactly 200 µL of 10 mM sodium nitroprusside prepared in phosphate buffer saline (pH 7.4) was mixed with 500 µL of the extract (or Trolox) at various concentrations of 0, 25, 50, 75 and 100 µL in an 8 x 12 deep-well plate. The plate was allowed to stand at room temperature of 30°C for 60 minutes. Then, 500 µL of the incubated mixture was added to 1.0 mL of sulfanilic acid reagent (prepared by mixing 33% sulfanilic acid in 20% glacial acetic acid) in a separate deep- well plate and incubated at room tempe- rature for 5 min. Then, 1.0 mL naphthylethy- lenediamine dihydrochloride (0.1% w/v) was mixed with the incubated mixture and allowed to stand at room temperature for additional 15 minutes. Finally, 200 µL of each mixture was transferred into 8 x 12 microplate and the absorbance was read at 540 nm was measured using a multi-well plate reader (Molecular Devices, Spectra- max Plus). The percentage reduction in NO accumulation was calculated as percen- tage of absorbance of wells with samples (A1) compared to wells without sample (A0) using this equation:
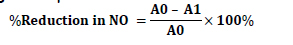
Chromatographic analysis identification of volatile components
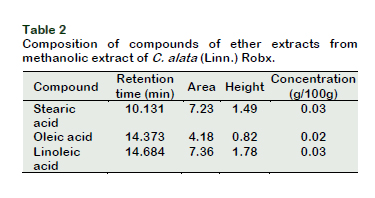
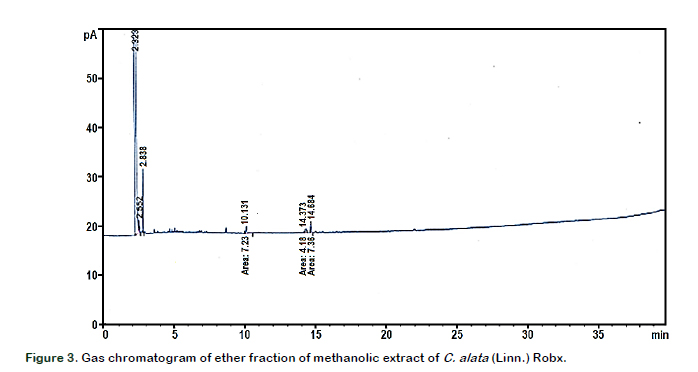
Gas chromatographic analysis was perfor- med using Agilent Technologies GC-FID (flame-ionization detection) (Model 7890) equipped with Agilent 19091S-433HP-5MS 5% Phenyl Methyl Silox column (30 m × 250 µm × film thickness 0.25 µm). Helium was used as carrier gas at a constant rate of 1.5 mL/min. The injector temperature was 250 °C. The concentrations of the identified fatty acids from the resulting chromato- gram were determined by the ratio of peak area of the internal standards and the area of the respective fatty acid.
Statistical analysis
The results were expressed as mean ± SD (standard deviation). Differences between the means were compared by one-way analysis of variance followed by Turkey- Kramer test for pairwise comparisons at p < 0.05
3. Results and discussion
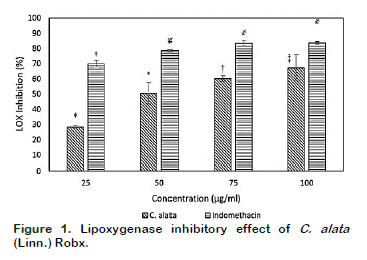
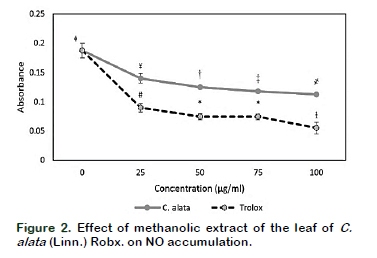
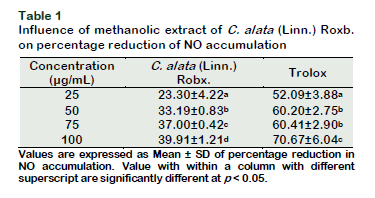
Results of the in vitro soyLOX inhibitory assay of the extract and the standard (indomethacin) are shown in Figure 1. The extract and the standard had noticeable effects on the percentage inhibition of soyLOX range of 28.39 ± 1.36 (at 25 µg/mL) to 67 ± 4.43 (at 100 µg/mL) and 69.86 ± 2.05 (at 25 µg/mL) to 83.73 ± 0.85 (at 100 µg/mL), respectively. Moreover, soyLOX inhibitory potential of the indomethacin solution only increased moderately but significant (p < 0.05) at tested concentrations of 75 and 100 µg/mL compared to the increase at 25 and 50 µg/mL. The study showed a similar pattern in the depletion of NO in NO accumulation assay (Figure 2). The extract and the standard significantly (p < 0.05) reduced the NO content at the tested concentrations. The methanolic extract at a concentration of 100 µg/mL had the highest influence in reducing the NO levels by up to 39.91 ± 1.21% (0.1127 ± 0.0022) compared to the control (0.1876 ± 0.0123). Moreover, at concentrations of 25, 50 and 75 µg/mL, the influence of the extract was corres- pondingly significant (p < 0.05), decreasing the NO accumulation by 23.30 ± 4.22% (0.1401 ± 0.0079), 33.19 ± 0.83% (0.1253 ± 0.0016) and 37.00 ± 0.42% (0.1182 ± 0.0008), respectively (Table 1). Moreover, trolox, which was used as the standard generated a considerable reduction in the NO levels by up to 70.67 ± 6.04% at 100 µg/mL compared to the control and decreased correspondingly at all tested concentration. The result of the GC/FID identification and quantification of volatile compounds is shown in Table 2 and the chromatogram is presented in Figure 3. The identified compounds comprise stearic, oleic and linoleic acids.
Many natural products of plant origin possess antioxidants that act in response to the endogenous generation of reactive oxidant species and other free radicals (Oso et al., 2018). There has been increasing interests recently in discovering natural products with medicinal benefits which can inhibit oxidative damage and prevent disorders that are related to inflammatory responses (Zeldin, 2002).
This study examined the soyLOX inhibitory activity of methanolic extract of C. alata (Linn.) Roxb. as well as its influence on the accumulation of NO. Excessive generations of metabolites of LOX such as leukotrienes play important roles in the progress of acute inflammation (Ivanov et al., 2010). LOXs could be inhibited by a number of chemicals which bind to or near the iron co- factor (Skrzypczak-Jankun et al., 2007). C. alata (Linn.) Roxb. has been used in traditional medicine in the treatment of pathologies that are associated with inflammatory disorders (Lewis and Levy, 2011). The methanolic extract of C. alata (Linn.) Roxb. showed the LOX inhibitory activity ranges from 33.69 ± 0.85 to 77.40 ± 8.61 at the concentrations range of 25µg/ml to 100µg/ml. The beneficial effect of the plant has been related to its contents of kaempferol, emodin, chrysoeriol, querce- tin, rhein, propelargonidins and phenols (Singh et al., 2012; Duong et al., 2013; Ramsay and Mueller-Harvey, 2016; Oso and Olowookere, 2018). The inhibition of soyLOX activity used in the present work could be presumed as predictive inhibition of the mammalian LOX system (Gundersen et al., 2003). The involvement of LOXs in progression of several pathologies has been supported by a number of literatures, suggesting that the inhibition of LOXs may denote promising therapeutic approaches in the regulation of arachidonic acid (Sarah et al., 2015). This result could validate the traditional use of the plant as remedy for inflammatory disorders. The structural features of plant polyphenols widely distri- buted in nature have been suggested to specially interfere with the LOX pathway (Hussain et al., 2005); thus, the anti- inflammatory properties of this extract could possibly be explained by the hypo- thetical inhibitory effects of its phenolic compounds on arachidonic acid metabo- lism. This is in agreement with previous claim that hydro-alcoholic extract of the botanical could exhibit anti allergic activity through inhibition of LOX (Singh et al., 2012). Nonetheless, the in vivo predis- position of the inhibitors might be influenced by the source of the enzyme, its isozyme and the oxidation state of co-factor (Pham et al., 1998; Borbulevych et al., 2004).
In addition, the potential of the extract to deplete NO was also investigated compa- rable to trolox, a standard antioxidant and an analogue of vitamin E. NO is a free radical and neurotransmitter with cyto- protective and cytotoxic action. It is usually formed as a product of metabolism of L- arginine by nitric oxide synthase. Despite the fact that its half-life in blood is short, due to its rapid clearance and inactivation by haemoglobin and myoglobin, NO can form complexes with other compounds that can serve as stable NO carriers thus enhancing its bioactivity (Pereira et al., 2011). Moreover, it has been indicated that bioactivities of NO can be potentiated as it can be generated non-enzymatically or enzymatically from nitrate and nitrite during tissue hypoxia or by disproportionation at low pH (Zweier et al., 1999; Li et al., 2008). The present study indicated that the methanolic extract of C. alata (Linn.) Roxb. considerably reduced the accumulation of NO in the reaction wells in concentration depended manner. This observation could substantiate previous reports on the antioxidant potential of the extracts of C. alata (Linn.) Roxb. (Chatterjee et al., 2013; Oso et al., 2018). NO has been extensively studied to explicate its role as a signalling molecule in metabolism, cell function and disease responses. NO reacts with mole- cules such as free radicals or transition metal ions that possess unpaired orbital electrons (Padmaja and Huie, 1993). Excessive generation of NO has been associated with various inflammatory disor- ders including cancer and cardiovascular diseases (Pereira et al., 2011). Decreasing the NO concentration during inflammatory responses might be helpful in lessening its threats in the interplay between cellular oxidative stress and inflammation.
Furthermore, based on the identification and quantification of volatile compounds present in the methanolic extract of the plant, it could be concluded that com- pounds such as stearic, oleic and linoleic acids could also contribute to the professed anti-inflammatory properties (Das, 2006; Carrillo et al., 2012).
Besides the suggested physiological roles of C. alata (Linn.) Roxb., its limiting actions on LOX and NO could also play important role in food processing. LOX have been associated with the generation of unde- sirable flavour and aroma during proces- sing and storage of proteinous plant pro- ducts (Robinson et al., 1995). Moreover, LOX may also lead to loss of essential nutri- ents and natural colorants by enhancing co-oxidation of carotenoids (Robinson et al., 1995). This suggests that C. alata (Linn.) Roxb. might also be used as a preservative agent to improve the storage quality of various food products.
4. Conclusions
In the present investigation, methanolic extract of C. alata (Linn.) Robx. was found to inhibit soyLOX activity in concentration- dependent manner and reduce accumula- tion of NO generated by sodium nitro- prusside. The observations show that the extract has the potential to interact with these biological targets. This perception on the inhibition of LOXs and clearance of NO could have shifting effects in the deve- lopment of anti-inflammatory drugs. Never- theless, additional works are encouraged to recommend its potential use as an alternative therapeutic agent against inflammatory disorders.
Acknowledgments
The assistance of Mr. Adebowale Olabanji of Bridge-Biotech Ltd, Ilorin, Nigeria is gratefully recognised.
References
Alitonou, G.A.; Avlessi, F.; Sohounhloue, D.K.; Agnaniet, H.; Bessiere, J.M.; Menut, C. 2006. Investigations on the essential oil of Cymbopogon giganteus from Benin for its potential use as an antiinflammatory agent. International Journal Aromatheraphy 16: 37- 41. [ Links ]
Borbulevych, O.Y.; Jankun, J.; Selman, S.; Skrzypezak-Jankun, E. 2004. Lypoxygenase interactions with natural flavonoid, quercetin, reveal a complex with protocatechuic acid in its X-Ray structure at 2.1 Å resolution. Proteins, Structure, Function and Bioinfor- matics 54:13-19. [ Links ]
Canals, S.; Casarejos, M.J.; de Bernardo, S.; Rodrı´guez-Martı´n, E.; Mena, M.A. 2003. Nitric Oxide Triggers the Toxicity due to Glutathione Depletion in Midbrain Cultures through 12-Lipoxygenase. Journal of Biolo- gical Chemistry 278 (24): 21542–21549. [ Links ]
Carrillo, C.; Cavia Mdel, M.; Alonso-Torre, S. 2012. Role of oleic acid inimmune system; mechanism of action; a review. Nutrición Hospitalaria 27(4): 987-990. [ Links ]
Chatterjee, S.; Chatterjee, S.; Deyand, K.K.; Dutta, S. 2013. Study of antioxidant activity and immune stimulating potency of the ethnomedicinal plant. Cassia alata (L.) Roxb. Medicinal and Aromatic Plants 2(4): 131. [ Links ]
Chua, L.Y.W.; Chua, B.L.; Figiel, A.; Chong, C.H.; Wojdylo, A.; Szumny, A.; Lech, K. 2019. Characterisation of the convective hot-air drying and vacuum microwave drying of Cassia alata: Antioxidant activity, essential oil volatile composition and quality studies. Molecules 24:1625. [ Links ]
Das, U.N. 2006. Essential fatty acids: bioche- mistry, physiology and pathology. Biotech- nology Journal 1:420-439. [ Links ]
Dennis, E.A.; Norris, P.C. 2015. Eicosanoids storm in infection and inflammation. Nature Reviews Immunology 15(8):511-523. [ Links ]
Duong, N.T.T.; Chinh, H.T.; Din, T.S.; Phong, T.L.H.; Phuong, P.N.; Quynh, P.Q.; Truc, N.T.T.; Quang, T.T. 2013. Contribution to the study on chemical constituents from the leaves of Cassia alata L. (Caesalpiniaceae). Science, Technology and Development 16(2): 26–31. [ Links ]
Garrat, D.C. 1964. The Quantitative Analysis of Drugs, Japan, Chapman and Hall. 456 pp. [ Links ]
Gundersen, L.L.; Malterud, K.E.; Negussie, A.H.; Rise, F.; Teklu, S.; Ostby, O.B. 2003. Indozilines as Novel Potent Inhibitors of 15- lipoxygenase. Bioorganic and Medicinal Chemistry 11: 5409–5415. [ Links ]
Hussain, T.; Gupta, S.; Adhami, V.M.; Mukhtar, H. 2005. Green tea constituent epigallocatechin- 3-gallate selectively inhibits COX-2 without affecting COX-1 expression in human prostate carcinoma cells, International Journal of Cancer 113: 660–669. [ Links ]
Ivanov, I.; Heydeck, D.; Hofheinz, K.; Roffeis, J.; O’Donnell, V.B.; Kühn, H. 2010. Molecular enzymology of lipoxygenases. Archive of Biochemistry and Biophysics 503: 161-174.
Khlifi, D.; Sghaier, R.M.; Laouni, D.; Hayouni, A.; Hamd, M.; Bouajila, J. 2013. Anti-Inflammatory and Acetylcholinesterase Inhibition Activities of Globularia alypum. Journal of Medical and Bioengineering 2(4):232-237. [ Links ]
Larsson, A.; Bӓck, M.; Hjoberg, J.; Dahlén, S. 2005. Inhibition of Nitric-Oxide Synthase Enhances Antigen-Induced Contractions and Increases Release of Cysteinyl-Leukotrienes in Guinea Pig Lung Parenchyma: Nitric Oxide as a Protective Factor. Journal of Phar- macology and Experimental Therapeutics 315 (1):458–465. [ Links ]
Leelaprakash, G.; Rose, J.C.; Dass, S.M. 2012. In vitro Anti-inflammatory activity of Momordica charantia by inhibition of lipoxygenase enzyme. International Journal of Pharmacy and Pharmaceutical Sciences 4 (S1): 148-152. [ Links ]
Lewis, A.; Levy, A. 2011. Anti-inflammatory activities of Cassia alata leaf extract in complete Freund’s adjuvant arthritis in rats. West Indian Medical Journal 60(6): 615–621.
Li, H.; Cui, H.; Kundu, T.K.; Alzawahra, W.; Zweier, J.L. 2008. Nitric oxide production from nitrite occurs primarily in tissues not in the blood: critical role of xanthine oxidase and aldehyde oxidase. Journal of Biological Chemistry 283: 17855-17863. [ Links ]
Li, J.; Rao, J.; Liu, Y.; Cao, Y.; Zhang, Y.; Zhang, Q.; Zhu, D. 2013. 15-Lipoxygenase promotes chronic hypoxia-induced pulmonary artery inflammation via positive interaction with nuclear factor-κB. Arteriosclerosis, Throm- bosis, and Vascular Biology 33: 971–979 [ Links ]
Medzhitov, R. 2008. Origin and physiological roles of inflammation. Nature 454: 428-435. [ Links ]
Oso, B.J.; Olowookere, B.D. 2018. A study on the total phenolics and antioxidant properties of different solvent extracts of dried leaves of Cassia alata (L.) Roxb. Medicinal Plants 10 (4): 348-352. [ Links ]
Oso, B.J.; Oyewo, E.B.; Oladiji, A.T. 2018. Phytochemical compositions, antioxidant capabilities and immunomodulatory in vitro indices of Xylopia aethiopica fruit extracts. Advance Pharmaceutical Journal 3(1): 29-37 [ Links ]
Padmaja, S.; Huie, R.E. 1993. The reactions of nitric oxide with organic peroxyl radical. Biochemical and Biophysical Research Communications 195:539–544. [ Links ]
Pereira, A.C.; Paulo, M.; Araújo, A.V.; Rodrigues, G.J.; Bendhack, L.M. 2011. Nitric oxide synthesis and biological functions of nitric oxide released from ruthenium compounds. Brazilian Journal of Medical and Biological Research 44(9): 947-957. [ Links ]
Pham, C.; Jankun, J.; Skrzypczak-Jankun, E.; Flowers, R.A.; Funk, M.O. 1998. Structural and thermochemical characterization of lipo- xygenase-catechol complexes. Biochemistry 37:17952-17957. [ Links ]
Rådmark, O.; Werz, O.; Steinhilber, D.; Samuelsson, B. 2015. 5-Lipoxygenase, a key enzyme for leukotriene biosynthesis in health and disease. Biochimica et Biophysica Acta 1851(4): 331–339. [ Links ]
Ramsay, A.; Mueller-Harvey, I. 2016. Senna alata leaves are a good source of propelargoni- dins. Natural Product Research 30(13):1548– 1551 [ Links ]
Robinson, D.S.; Wu, Z.C.; Domoney, C.; Casey, R. 1995. Lipoxygenases and the quality of foods. Food Chemistry. 54: 33–43. [ Links ]
Sarah, A.; Tersey, E.B.; Theodore, R.H.; David, J.M.; Jerry, L.N.; Raghavendra, G.M. 2015. Minireview: 12-Lipoxygenase and Islet β-Cell Dysfunction in Diabetes. Molecular Endo- crinology 29: 791– 800. [ Links ]
Serhan, C. N. 2002. Endogenous chemical mediators in anti-inflammation and pro- resolution. Current Medicinal Chemistry 1: 177-192. [ Links ]
Shinde, U.A.; Kulkarni, K.R.; Phadke, A.S.; Nair, A.M.; Mungantiwar, A.A.; Dikshit, V.J.; Saraf, M.N. 1999. Mast cell stabilizing and lipoxygenase inhibitory activity of Cedrus deodara (Roxb.) Loud. Wood Oil. Indian Journal of Experimental Biology 37(3): 258- 261. [ Links ]
Singh, B.; Nadkarni, J.R.; Vishwakarma, R.A.; Bharate, S.B.; Nivsarkar, M.; Anandjiwala S. 2012. The hydroalcoholic extract of Cassia alata (Linn.) leaves and its major compound rhein exhibits antiallergic activity via mast cell stabilization and lipoxygenase inhibition. Journal of Ethnopharmacology 141(1): 469- 473. [ Links ]
Skrzypczak-Jankun, E.; Chorostowska-Wynimko, J.; Selman, S.H.; Jankun, J. 2007. Lipoxy- genases – A challenging problem in enzyme inhibition and drug development. Current Enzyme Inhibition 3: 119-132. [ Links ]
Wisastra, R.; Dekker, F.J. 2014. Inflammation, Cancer and Oxidative Lipoxygenase Activity Are Intimately Linked. Cancers 6:1500-1521. [ Links ]
Zeldin, D.C. 2002. The 5-Lipoxygenase Pathway: A New Therapeutic Target for the Treatment of Pulmonary Fibrosis. American Journal of Respiratory and Critical Care Medicine 165: 146-147. [ Links ]
Zweier, J.L.; Samouilov, A.; Kuppusamy, P. 1999. Non-enzymatic nitric oxide synthesis in biological systems. Biochimica Biophysica Acta 1411: 250-262. [ Links ]
Corresponding author
E-mail: basjoe08@gmail.com (B.J. Oso)
Received February 11, 2019.
Accepted March 17, 2019.

