










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkScientia Agropecuaria
versão impressa ISSN 2077-9917
Scientia Agropecuaria vol.10 no.2 Trujillo abr./jun. 2019
http://dx.doi.org/10.17268/sci.agropecu.2019.02.08
ARTÍCULOS ORIGINALES
Vermicompost enriquecido con microorganismos benéficos bajo dos sistemas de producción y sus efectos en el rábano (Raphanus sativus L.)
Microbial enrichment vermicompost under two production system and its effects on radish (Raphanus sativus L.) production
Silvia Velecela1,2,*; Victor Meza2; Sady García2 ORCID iD https://orcid.org/0000-0002-2498-3940; Julio Alegre2 ORCID iD https://orcid.org/0000-0002-7282-045X; Carlos Salas3 ORCID iD https://orcid.org/0000-0002-1641-1571
1 Universidad Católica de Cuenca – UCACUE, Avenida de las Américas y Humbolt, Cuenca, Ecuador.
2 Universidad Nacional Agraria La Molina – UNALM, Av. La Molina s/n, Lima, Perú.
3 Universidad Técnica de ManabíUTM, Avenida José María Urbina s/n Portoviejo, Ecuador.
Resumen
Se evaluó la calidad de vermicompost resultante de excreta de vacuno bajo dos sistemas de producción a través de sus parámetros físico químicos y de su efecto sobre el crecimiento y el rendimiento de plantas de rábano (Rhaphanus sativus L) bajo condiciones de campo en la Universidad Nacional Agraria la Molina (UNALM), Lima, Perú. Se empleó un diseño de bloques completo al azar (DBCA) 12 tratamientos con tres repeticiones. El primer factor de estudio incluyó dos tipos de compost de vacuno, compostados bajo dos tratamientos (lavado e inoculado con microorganismos benéficos). Como segundo factor se empleó dos tipos de microorganismos (Bacillus sp., y microorganismos benéficos). Un tercer factor incluyó dos sistemas de producción de vermicompost (zanja y techo a dos aguas) empleando Eisenia foetida (40 días). Los vermicomposts procedentes del sistema zanja presentaron mayor descenso en salinidad, pH, relación C/N, contenido total de fósforo, potasio, sodio y mayor incremento en contenido de humedad, calcio, magnesio y sustancias húmicas. C/N indicó la estabilización del vermicompost. En el rendimiento del rábano el tratamiento MZM (compost inoculado con microorganismos benéficos del sistema zanja) alcanzó mayor altura y peso fresco de hojas y peso de hipocótilo.
Palabras clave: estiércol vacuno; microorganismos benéficos; vermicompost; Eisenia foetida.
Abstract
Vermicompost production from livestock manure was evaluated under two production systems through their chemical parameters and evaluating its effect on the growth and yield of radish (Rhaphanus sativus L.) under field condition at the National Agrarian University La Molina, in Li ma – Perú. The experimental design was a complete randomized block design (CRBD) with 12 treatments and three replications. The first factor of study includes two different types of livestock manure that was pre-decomposed under two treatments (washing and inoculation with beneficial microorganism consortium). As second factor, included two kinds of microorganism (Bacillus sp., and beneficial microorganism’s consortium). The third factor included two vermicomposting production system (superficial named "gable roof" and underground "pit system’’) processed with Eisenia foetida during 40 days. The characterization of vermicomposts from the pit system presented greater reduction in salinity, pH, C/N, total phosphorus, potassium, sodium concentration, higher retained moisture, total calcium, magnesium and humic subtances concentration. The C/N value showed that the vermicompost was stable. By comparison, the means indicate that treatment MMZ (livestock manure pre-decomposed inoculate with beneficial microorganism under pit system) attained greater height and length of leaves and individual weight of fruit.
Keywords: livestock manure; beneficial consortium; vermicompost; Eisenia foetida.
1. Introducción
El fuerte crecimiento demográfico constante ha impulsado en gran parte el incremento de la demanda mundial del consumo de carne de vacuno. Se prevé que para 2026 aumentará alrededor del 6% en los países desarrollados, mientras que en las regiones en desarrollo, probablemente se incrementará al 17% (OCDE-FAO, 2017). Conjuntamente a esta intensificación; el consumo y la demanda de mercado ha incrementado la generación de los residuos biológicos agropecuarios, cuya inadecuada gestión provoca contaminación del aire, del agua superficial y del suelo (Wang et al., 2015). A pesar de que las características del estiércol están influenciadas por factores, como la alimentación y la edad del animal, generalmente se asume que la cantidad de nutrientes eliminados en las excretas es proporcional a la que toman en su alimentación (Diacono y Montemurro, 2010). El estiércol vacuno presenta gran cantidad de elementos como nitrógeno, fósforo y potasio y debido a la falta de gestión en su manejo y tratamiento se han convertido en un importante contaminante ambiental; por su disposición directa sobre el suelo y a cielo abierto (Pinos-Rodríguez et al., 2012; Sifolo et al., 2018). El compostaje y vermicompostaje han sido propuestos como tecnologías de manejo integrado; métodos limpios y sostenibles para aprovechar y reducir el volumen de residuos, como una opción potencial para su bio oxidación, degradación y estabilización, acelerando el procesos de descomposición de dos a cinco veces (Bhat et al., 2016; Ali et al., 2018). En los últimos 30 años, la vermicultura ha captado considerable atención (Wani et al., 2013) y progresivamente ha alcanzado relevancia en Europa, África, Asia, Australia y América Latina, donde su desarrollo ha impactado ya como una realidad productiva y rentable en el mercado a mediano plazo (Schuldt, 2006; Balasubramani et al., 2019), las lombrices reducen el volumen de los residuos orgánicos que ingieren en un 40 a 60%, produciendo deyecciones diarias aproximadas a su peso corporal y equivalentes al 50% de lo que ingieren (Nagavallemma et al., 2006). Aliviando los problemas ambientales ante la generación de miles de toneladas de estiércol, alta DBO (demanda biológica de oxígeno), pérdida de nutrientes, salinidad, elevado pH entre otros difíciles de remediar (Mitchell, 1997). Simultáneamente, al incrementar su biomasa son empleadas como fuente de proteína o suplemento de dieta animal; produciéndose un alimento a partir de residuos orgánicos (Kostecka y Paczka, 2006) y a la vez un fertilizante orgánico (Ali et al., 2018); dos productos valiosos dentro del mercado y sostenibles para el medio ambiente (Lim et al., 2016; Nurhidayati et al., 2018). En 1971, Teruo Higa de la Universidad de Ryukyus, Japón introdujo el uso de microorganismos eficientes (EM®) (Higa y Parr, 1989). Desde entonces, diferentes inóculos han sido probados para promover el crecimiento de plantas a través de la promoción de rizobacterias, microorganismos beneficiosos para el crecimiento vegetal, producción y calidad de cultivo, entre ellos: Alcaligenes sp., Arthrobacter sp., Azotobacter sp., Pseudomonas sp. (Amini et al., 2015; Mahmud et al., 2018). Sharma et al. (2017) inoculando un consorcio de (EM) Candida tropicalis (Y6), Lactobacillus sp., Streptomyces globisporus (C3) y bacterias fotosintéticas incrementaron la actividad microbiana y el contenido de carbón orgánico y nitrógeno disponible, mejorando la fertilidad del suelo. Se ha demostrado, que el proceso de vermicompost es acelerado empleando la técnica de enriquecimiento microbiano, partiendo de esta combinación se obtiene un producto final enriquecido que presenta múltiples benéficos del conglomerado de microorganismos residentes en el producto final, en base al componente microbiano y a su actividad influyendo en la fijación de nitrógeno, absorción de nutrientes principales, control o supresión de enfermedades (Gupta, 2012; Olle, 2018). Das et al. (2016) verificaron el aumento significativo de la población microbiana y actividad enzimática aplicando consorcios de Azotobacter chroococcum (fijador de nitrógeno), Bacillus firmus (solubilizador de potasio) y Paenibacillus polymyxa (solubilizador de fosfato). Kumar et al. (2011) obtuvieron resultados similares con Aspergillus niger, Pleurotus sajor-caju y Trichoderma viride. Otra alternativa biológica planteada es la suplementación con microorganismos probióticos como Lactobacillus sporogenes, Saccharomyces cerevisiae (Saravana y Aruna, 2013) aportando con una dieta más exigente. De los grupos microbianos se ha puesto énfasis en investigar el verdadero valor de las especies de Bacillus sp., por sus grandes beneficios en el sector de la agricultura inhibiendo patógenos de plantas. Por ejemplo; Bacillus subtilis ENT2 secreta fengicina, un biosurfactante que exhibe gran actividad antifúngica (Sumi et al., 2015), al mismo tiempo generan esporas latentes resistentes a la radiación, temperaturas, degradación enzimática, etc. (Lee et al., 2012). El objetivo del estudio fue evaluar la influencia de la técnica de inoculación microbiana empleando microorganismo benéficos y cepas de Bacillus sp., en el proceso de vermicompostaje para acelerar la degradación de la excreta pretratada pura de vacuno empleando dos diferentes sistemas de producción que permitan generar información sistematizada de las características físico químicas del producto obtenido evaluando sus efectos sobre el rendimiento de cultivo del Raphanus sativus L, como una opción más prometedora como reconocimiento global de la necesidad de recuperar materiales orgánicos útiles, ampliando el conocimiento ante la limitada información que existe sobre los beneficios del enriquecimiento microbiano.
2. Materiales y métodos
Inóculo microbiano
El consorcio de microorganismos benéficos y las cepas de Bacillus sp., fueron donados por el Laboratorio de Biología de la UNALM, el consorcio se obtuvo de hojas de plantas. El inóculo de Bacillus sp., se cultivó en caldo nutritivo a 25 ºC. Se empleó por cada kg de sustrato 50 mL de inóculo con una concentración de 35x108 UFC por mL, un volumen igual se empleó del consorcio de microorganismos benéficos para cada tratamiento.
Estiércol vacuno
El estiércol vacuno se obtuvo del establo de la Universidad Agraria, La Molina, tras un proceso combinado fue pretratado seis semanas (lavado e inoculado) y posteriormente compostado durante ocho semanas previo al proceso de vermicompostaje, las características químicas de los compost resultantes de los dos tratamientos aplicados se presentan en la Tabla 2.
Vermicompostaje
Se evaluaron dos sistemas de producción de vermicompostaje durante 40 días. Para el sistema de zanja se cavaron zanjas rectangulares (1 m de largo x 1 m de ancho x 0,40 m de altura) y para el sistema techo a dos aguas sobre la superficie se edificaron estructuras (1 m de largo x 1m de ancho x 0,40 m de altura). Se empleó como sustrato 20 kg de compost lavado e inoculado con microorganismos benéficos, aplicando por kg 50 mL, tanto del consorcio de microorganismos benéficos y Bacillus sp. El porcentaje de humedad se mantuvo alrededor del 70% de la máxima capacidad de retención mediante riego frecuente empleando una cinta microperforada de aspersión (tipo Santeno®, I) con las siguientes características: diámetro interno de 28 mm, espesor de cinta de 0,24 mm y puntos de emisión espaciados cada 0,15 m. Cada cinta de aspersión se instaló en el medio de cada bloque experimental. Se inocularon 1000 lombrices del tipo (Eisenia foetida) procedentes del pie de cría de la unidad experimental de vermicultura de la UNALM., dentro de cada tratamiento en sus respectivos sistemas. Se seleccionó individuos juveniles, teniendo en cuenta el tamaño y la presencia de un anillo clitelar desarrollado y visible, lo cual indica la capacidad reproductiva del organismo (Schuldt et al., 2005). El vermicompost fue muestreado cada ocho días tomándose muestras compuestas de aproximadamente 300 g.
Bioensayo
Se empleó una maceta con capacidad de 5 kg, para el sustrato se empleó 2 kg de vermicompost resultante de cada unidad experimental con 2 kg arena previamente lavada de conductividad 0,47 dS m-1, sembrado con cinco semillas de Raphanus Sativus L. (Tabla 1).
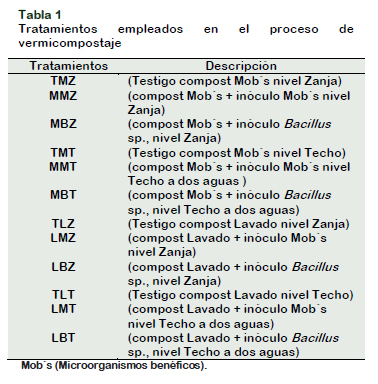
Métodos
En el Laboratorio de suelos de la UNALM, se caracterizaron las muestras cada 8 días para: % de humedad, EC, pH, tanto al inicio y al término del proceso de vermicompostaje, mientras que N, P205, K2O, Mg, Na y sustancias húmicas se realizaron final del proceso. El pH y C.E del vermicompost fueron determinados en agua destilada relación 1:10 (p/v), empleando el potenciómetro-conductímetro Consort® C1020. El contenido de humedad fue determinado por secado a peso constante empleando la estufa (Venticell LSIS-B2V / VC 222) a 75 ºC. La temperatura se usó un termómetro digital. El nitrógeno total de Kjeldahl (TKN) se estimó pesando 0,25 g mediante micro-Kjeldahl, y el carbono orgánico total (TOC) se midió mediante calcinación introduciendo la muestra durante 5 h a 550 ºC, en la mufla Barnstead Thermolyne 62700. En el proceso de digestión húmeda para determinar P2O5, se utilizó como extractante, NaHCO3, con una solución sulfo-molibdica. Desarrollado el color, se registró lecturas de absorbancia en el espectro fotómetro modelo Helios Epsilon, a 660 nm de longitud de onda. K2O, CaO, MgO y Na se determinaron con el extracto anterior en un equipo de absorción atómica (Perkin Elmer® AAnalyst 200). Los ácidos húmicos y fúlvicos se determinaron por el método de fraccionamiento (Black et al., 1965), 2 g de la muestra secada a 100 ° C durante 24 horas en un horno (modelo BLUE M MW-17TC), la extracción se realizó con una mezcla de 0,1 M de pirofosfato agregando NaOH 0,1 M, agitando durante una hora y centrifugando durante 25 minutos a 4500 rpm con un ordenador (IEC Size 2 Model K), secado el precipitado se determinó el % huminas. Se aplicó un Diseño de Bloques Completo al Azar (DBCA), con 12 tratamientos y 3 repeticiones, originando 36 unidades experimentales de 3 bloques en campo, cada sistema contó con un tratamiento testigo sin inoculación microbiana para ambos sistemas, para la evaluación del Raphanus Sativus L. se empleó los vermicomposts resultantes, se evaluó la altura de planta a los 15 y 30 días de siembra y 45 días de cosecha empelando una regla. Al término de la cosecha se determinó el peso del fruto en g empleando la balanza digital (Sartorius CP323 S).
Para la evaluación estadística se usó el programa estadístico InfoStat/E con especificación del modelo en R para determinar las variables con sus respectivas diferencias estadísticas empleando la prueba de hipótesis marginal LSD Fisher (Alfa=0,05) del Modelo Lineal General Mixto.
3. Resultados y discusión
3.1. Caracterización de vermicompost resultante en los dos sistemas
La caracterización físico química de los compost y vermicompost (40 días), se indica en la (Tabla 2). Durante este tiempo, se controló el porcentaje de humedad el cual fue apropiado para el desarrollo de las lombrices, oscilando entre un 54,22 en el sistema zanja y 66,21% en el sistema techo (p-value 0,0389), mayor porcentaje de humedad se registró en el tratamiento LBZ con un 66,8 % mientras que el menor en el tratamiento TLT 51,65 %. Esta tendencia de diferencia entre ambos sistemas puede deberse a que una gran fracción de la humedad del sustrato en el sistema techo a dos aguas podría perderse de igual manera como sucede en el suelo y que ha sido descrito por Wang et al. (2012) a través de la evaporación directa, del incremento de difusión de vapor, así como también debido a mayores temperaturas registradas en este sistema, propiciadas por la influencia que ejerce la cubierta del sistema sobre substrato a manera de techo, en contraste con el sistema de zanja en donde la unidad experimental es de mayor dimensión lo que permite mejor dispersión de la temperatura. Se evidenció que existe mayor infiltración en el sistema superficial debido a que el agua fluye con mayor velocidad de un nivel alto a un bajo por la fuerza de la gravedad como escorrentía superficial, mientras que un flujo de agua subterránea (sistema zanja) debido a su gradiente inferior es más lento que el escurrimiento superficial (Chidavaenzi et al., 1997), esta podría ser una de las razones por las cuales el sistema zanja retiene mayor cantidad de agua. Los valores obtenidos son considerados como normales, debido a que el vermicompostaje requiere una humedad entre 70 a 90 %, contrario al proceso de compostaje que requiere entre 40 hasta 60 %. Fernández-Gómez et al. (2010) indican que Eisenia foetida sobrevive entre 50 hasta 90 % de humedad, Wani et al. (2013) concuerdan de igual manera que un rango óptimo de humedad oscila entre el 60 y 90 %. Se debe considerar que el contenido de humedad es un factor crucial que afectaría la emisión de gases como amoníaco y metano durante el vermicompostaje, al igual que podría afectar las características nutrimentales, estructura física, porosidad, aireación, drenaje a lo largo del proceso (Lim et al., 2016).La conductividad eléctrica (CE) disminuyó en todos los tratamientos y entre los 2 sistemas se aprecia un patrón similar de disminución. En el transcurso del proceso se observó aumentó de CE entre la semana tres y cuatro, lo cual podría estar relacionada con la liberación de diferentes iones, como fosfato, amonio, potasio, etc. (Majlessi et al., 2012; Natchimuthu et al., 2019). Valores menores de CE se obtuvieron en el tratamiento MMZ el cual disminuyó partir de 5,79 a 2,45 dSm-1, mientras que en el tratamiento LBZ decreció a partir de 4,88 dSm-1 a 2,53 dSm-1, al contrario, el valor más alto 3,46 dSm-1 se registró en el tratamiento TLT que no recibió ningún tipo de inóculo, seguido de los tratamientos LBT 3,45 dSm-1 y LMT 3,43 dSm-1. Los valores de conductividad obtenidos son similares a los reportados por Mitchell (1997) al emplear 3 diferentes tipos de lechos: horizontal (1,5 m x 1,5 m x 0,3 m), surco (3 m x 1,5 m x 0,25 m) y vertical (1,5 m x 1,5 m x 0,2 m), con 0,09 m3 ; 019 m3 y 0,225 m3 de estiércol vacuno, registraron valores de 2,5 ; 3,1 y 3 dSm-1. Valores similares fueron registrados por Mousavi et al. (2017) de 2,32 dS m-1 a partir de una mezcla de residuos orgánicos con estiércol vacuno, mientras que Lazcano et al. (2008); Elvira et al. (1998) registrando valores cercanos 3 dS m-1. Para Hernández et al., (2011) el contenido de sales solubles va disminuyendo a medida que se van irrigando los lechos de vermicompostaje para mantener el porcentaje óptimo de humedad por efecto de lixiviación, acompañado de menor producción de metabolitos como amonio NH4+, así como también disminución de la precipitación de las sales disueltas resultando en la obtención de valores inferiores en los valores de CE (Lazcano et al., 2008). En el análisis de pH se encontraron rangos más bajos que los medidos en el sustrato inicial (8,32). Estos valores se asemejan a los hallazgos realizados por otros investigadores (Elvira et al., 1998; Suthar, 2009; Bhat et al., 2016). Cabe resaltar que no se encontraron diferencias significativas entre los tratamientos para los niveles de pH (pvalue 0,1614). No obstante, la máxima reducción de pH se encontró en el tratamiento LBT 7,70 seguido del tratamiento TMT 7,80. Este cambio en el pH puede deberse a la descomposición microbiana (Suthar y Singh, 2008a). Ndegwa et al. (2000) argumentan que su cambio podría ser atribuido al proceso de mineralización del nitrógeno, como a la del fósforo en nitritos / nitratos y ortofosfatos, la bioconversión de la materia orgánica en especies intermedias de los ácidos orgánicos o también ligado al crecimiento de la biomasa de las lombrices. Suthar (2009) resalta que la producción de CO2, amoníaco y descomposición microbiana son variables que pueden intervenir en la reducción de los niveles iniciales de pH del sustrato. El contenido de carbono total de un vermicompost, está relacionado con el contenido de materia orgánica. Edwards et al. (2011) indican que el carbono es un indicador importante de la estabilización de la materia, este disminuyó significativamente (p-value 0,0096) en todos los tratamientos durante el proceso de vermicompostaje. En el tratamiento MMZ disminuyó a 16,74%, seguido por el tratamiento MRM 16,91%, mientras que en los tratamientos con compost lavado en el tratamiento LBZ se registró 16,79%. Por el contrario, Sultana et al. (2015) reportaron un porcentaje mayor de 17,8%, también medido por método de ignición, esta disminución puede deberse al potencial combinado de acción que ejercen los microorganismos inoculados conjuntamente con la acción de las lombrices para desintegrar el material ingerido (Patidar et al., 2013). El nitrógeno total disminuyó reflejando valores 1,7 y 1,54% con un comportamiento muy similar entre los tratamientos y sus sistemas, no se encontraron diferencias significativas (p-value 0,7043). La tendencia ascendente, exhibida a lo largo de este proceso, es la resultante del consumo de carbono y a su vez a la combinaci&oa cute;n de las variables de temperatura y pH. Sin embargo, la disminución del pH puede ser un factor importante en la retención de nitrógeno debido a que este elemento se pierde como amoníaco volátil a valores altos de pH (Kaviraj y Sharma, 2003).
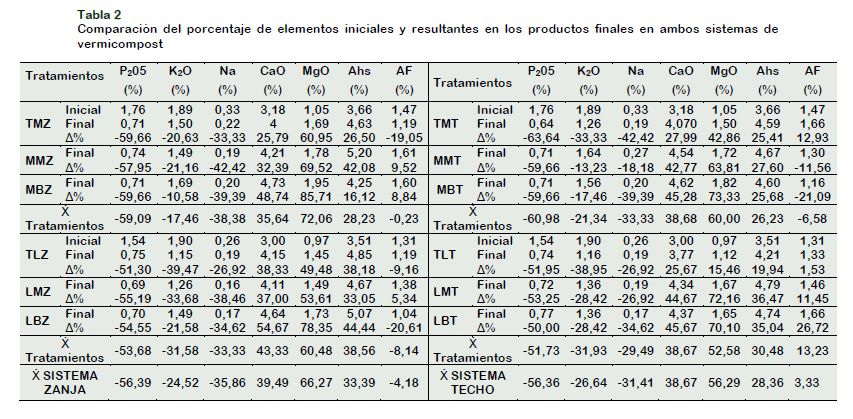
Akbar Babael et al. (2016) reportan que hay una conversión de este elemento a lo largo de su crecimiento disponible como proteína para la lombriz, obteniendo menor contenido de nitrógeno en el producto final. A través de esta experimentación, se ha demostrado que es posible la estabilización de la materia orgánica a través del proceso combinado de compostaje y vermicompostaje en sólo 40 días de estiércol de vacuno, sin adición de material extra. Al final del proceso las relaciones C/N fluctuaron entre valores de 10 y 12 similar al valor de 10,42 registrado por Vázquez y Loli (2018), habiendo partido de una relación inicial C/N de 22,5 e inferior al índice inicial 25 a 30 propuesto por Edwards et al. (2011) y diferente al de 50 experimentado por Lim et al. (2015), quienes al mezclar racimos de fruta con estiércol de caballo obtuvieron un producto final con una relación C/N igual 20. Esta reducción se debe también a que las lombrices liberan una parte del carbono como CO2 en el proceso de respiración, producción de moco y junto a sus deyecciones nitrogenadas (Barillari et al., 2006; Tripathi y Bhardwaj, 2004). La mutua acción de su sistema digestivo conjuntamente a las enzimas producidas por los microorganismos aceleran la degradación de la materia orgánica compleja, al igual que la producción de hormonas para su crecimiento (Karsten y Drake, 1995). En relación a los nutrientes presentes en el vermicompost, ciertos elementos exhiben un incremento, resultante de la mineralización de la materia orgánica, la acción de enzimas secretadas en el moco de la lombriz (fosfatasa), las cuales mejoran la degradación de la materia orgánica y la liberación de los metabolitos disponibles, e incluso el tipo de dieta alimenticia que recibe la lombriz (Das et al., 2016).
En la Tabla 2 se detallan los rendimientos obtenidos dentro de cada sistema y en cada tratamiento. Los datos revelan que el fósforo total (% PT) disminuyó en los vermicomposts resultantes comparado con el porcentaje inicial. A pesar de ello, se encontró incremento significativo entre los tratamientos (p-value 0,01). A nivel de zanja y techo los tratamientos con mayor (% PT) fueron el tratamiento TLZ 0,75 y LBT 0,77%, mientras los de menor porcentaje fueron LMZ 0,69 y TLT 0,64%. Al analizar la pérdida total por sistema se obtuvo un valor similar del 56,37% en ambos (Tabla 2). La liberación de P es atribuida a la acción que ejercen los microorganismos solubilizadores de fósforo presentes en los sustratos, así como a la actividad de las lombrices, mientras que su pérdida podría deberse a la lixiviación a lo largo del proceso. Suthar y Singh (2008b), indicaron que el contenido de P del abono orgánico puede incrementar por acción de las fosfatasas intestinales de las lombrices, a través de algunos estudios se ha identificado que las lombrices segregan fosfatasa alcalina (fosfatasa monosterasa) en sus deyecciones, la cual está a su vez involucrada en el ciclo P (Aira et al., 2007).
Se encontraron diferencias significativas (p-value 0,0104), al cuantificar la pérdida de %TK entre el sistema zanja y techo a dos aguas, entre sus tratamientos con compost inoculado estos exhibieron pérdidas de alrededor de (10,58 21,16%) y (13,23 33,33%) mientras que los tratamientos con compost lavado registraron valores entre (21,58 39,47%) y (28,42 38,95%). En el sistema zanja MBZ mostró el contenido más elevado de (% KT) 1,69%, mientras que el mínimo el tratamiento TLZ 1,15%. En el sistema techo a dos aguas el máximo incremento se cuantificó en MMT 1,64% y el mínimo en TLT 1,16%. Los resultados de pérdida total de los tratamientos en los sistemas describen una disminución de 24,52% en el sistema zanja y 26,64% en el de techo a dos aguas. Al compararlos los valores con los resultados presentados por Wani et al. (2013) de 0,88 % en compost de vacuno sin inoculación microbiana, discrepan debido a que %TK encontrado fue de (1,69%). Este hallazgo confirma la asociación existente entre la acción desempeñada por la carga microbiana inoculada conjunta a la actividad digestiva de la lombriz en la solubilización de ese elemento, teniendo en cuenta la fuente de nutrición empleada (Tabla 2). Además, Kaviraj y Sharma (2003) señalan que la microflora de las lombrices puede influir en el nivel de micronutrientes disponible, entre ellos el potasio, debido a la produc-ción de ácidos como el carbónico, nítrico y sulfúrico por los microorganismos, convir-tiéndose en un mecanismo importante en la solubilización del K insoluble.
Al inocular los tratamientos se promueve el aumento de la microflora presente en el intestino de las lombrices, y a su vez como lo han descrito otros investigadores aumenta la producción de ácidos incrementando el porcentaje de este elemento como se aprecia en los tratamientos MBZ (1,69%), LBZ (1,49%), MMT (1,64%), LBT (1,36%) en contraste al valor encontrado en los no inoculados. Esta asunción concuerda con lo indicado por Das et al. (2016) quienes sostiene que la producción de ácido por parte de los microorganismos es el principal mecanismo de solubilización no solo de fósforo sino también de potasio insoluble. Un considerable número de investigadores dependiendo del tipo de substrato han registrado mayores cuantificaciones (Akbar Babael et al., 2016; Das et al., 2016; Maji et al., 2017; Singh y Sharma, 2002). Se ha demostrado que es posible incrementar el contenido de este elemento empleando bacterias como Acidothiobacillus ferrooxidans, Bacillus mucilaginosus, Bacillus edaphicus, Bacillus circulan y Paenibacillus, las cuales han demostrado su gran capacidad para solubilizar el K con eficacia transformándolo en formas mayormente disponibles (Etesami et al., 2017). De los resultados, mayor disminución del porcentaje de sodio total (% NaT) (p-value 0,0044) se identificó en el sistema zanja en los vermicompost producidos con compost inoculado, el catión fue lixiviado en un 35,86%. MMZ exhibió la mayor pérdida (0,19%), mientras que en los vermicompost con compost lavado su porcentaje se redujo en un (0,16%) valor identificado en tratamiento LMZ. Conseguir la reducción de este catión es importante puesto que podría afectar el desarrollo, crecimiento y reproducción de las lombrices. Hughes et al. (2009) indicó que Eisenia foetida es sensible a la concentración de 131,58 mmol/kg de NaCl sustentando este valor tras aplicar la prueba de toxicidad (LC50), además registró que esta concentración afectó la puesta de cocones, el peso de lombriz y ocasionó estrés en las lombrices adultas. No obstante, un factor que podría contribuir con la reducción de la concentración de sales es el lavado con niveles de agua tolerables para retener la mayor cantidad de nutrientes beneficiosos de la planta, de esta manera la inoculación microbiana de microorganismos benéficos promovería su disolución puesto que se aprecia menor concentración en los tratamientos que fueron inoculados (Tabla 2). En relación al porcentaje total de calcio (% CaT) se cuantificó un incremento significativo durante todo el proceso de vermicompostaje. A pesar de que no se registraron diferencias significativas (p-value 0,7062).
El incremento total resultante fue de 35,86 en los tratamientos del sistema zanja y 31,34% en los del sistema techo a dos aguas. En los productos finales, se cuantificó la mayor cantidad de calcio en el tratamiento MMZ 4,73%, MBT 4,62%, LBZ 4,64%, LBT4,37% (Tabla 2).
En cuanto al incremento estudios anteriores han demostrado que este elemento es producido por las lombrices en sus glándulas calcíferas junto al carbonato de calcio y que a su vez es secretado en su mucosa, pues tienden a eliminar su exceso de producción para regular su pH intestinal y los fluidos celómicos, razón por la cual incrementa consecuentemente el contenido de este macronutriente en cada una de sus deyecciones y en las excreciones producidas por su cutícula. Según Karaca (2011) indica que las deyecciones frescas contienen altas concentraciones solubles en sal de este catión. Otro factor explicado por Sierra et al. (2013) podría deberse al efecto de concentración y a la conductividad eléctrica. No obstante, su contenido podría promover la precipitación de P liberado durante la degradación de la materia orgánica y al mismo tiempo evitar la lixiviación de ambos nutrientes. Con respecto a las concentraciones de magnesio total (% MgT) aumentaron en relación al inicio del proceso, lo que podría deberse a la actividad de las lombrices conjuntamente con los microorganismos, exhibiendo diferencias significativas (pvalue 0,0318). El mayor incremento 66,27% ocurrió en el sistema zanja, en contraste al sistema techo a dos aguas en el cual se registró un 56,29%. El tratamiento MBZ incrementó su porcentaje de 1,05% a 1,95% describiendo un aumento máximo de (85,71%), mientras que LBZ produjo un incremento del (78,35%). Por otro lado, menor porcentaje ocurrió en TLT (5,46%), Tabla 2.
Al termino del proceso se constató la humificación de los vermicompost registrando mayor contenido porcentual de ácidos húmicos (33,39%) en el sistema zanja y 28,36% en el sistema superficial techo a dos aguas. En MMZ el incremento fue de 3,66% valor cuantificado en el compost antes del proceso a 5,2% en el producto final, mientras que en LBZ incremento a partir de 3,51 a 5,07%. Los ácidos fúlvicos incrementaron en porcentaje, mientras que las huminas registraron una disminución, debido a la mayor mineralización de la materia (Tabla 2).
3.2. Influencia del vermicompost en el desarrollo de Raphanus sativus
3.2.1. Altura de plantas
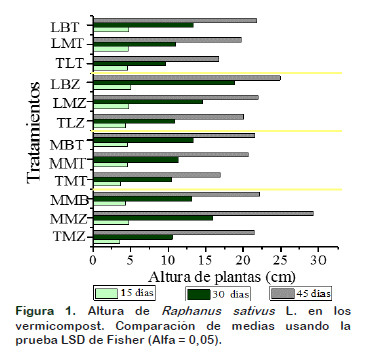
El efecto de los vermicompost en el crecimiento de las plantas no fue homogéneo. A los 15, 30 y 45 días de cosecha, se encontraron diferencias significativas (Figura 1). La altura de plantas varió de 3,62 cm a 5,06 cm a los 15 días, mayor media en altura se encontró en el tratamiento LBZ, mientras que la menor media en altura se registró en el tratamiento TMZ (p-value de 0,029). A los 30 días, la mayor media con una altura de 18,88 cm se encontró en LBZ y la menor altura con una media de 9,67 cm en el tratamiento TLT registrándose diferencias significativas entre los tratamientos (pvalue 0,0001). A los 45 días, el tratamiento MMZ tiene la presentó la mayor media 29,34 cm, en tanto que el tratamiento TLT exhibió la menor media con una altura de 16,77 cm con un (p-value 0,0001). La C.E en los tratamientos varió de 2,45 a 3,46 dSm-1. Ghosh et al. (2014) señalan que diferentes niveles de salinidad podrían afectar el crecimiento de las plantas y lo verificó al probar tres variedades de rábano encontraron que los diferentes niveles de salinidad afectaron el crecimiento de las plantas registrando un valor de 29,02 cm a una C.E de 4 dSm-1. Para Atiyeh et al. (2000) las diferencias entre crecimiento de las plantas no pueden únicamente deberse a su contenido nutricional o a factores físico químicos, sino también a que probablemente hay insumos que están posiblemente relacionados con el crecimiento de las plantas como son las sustancias biológicas originadas por los microorganismos, los cuales según el reporte de Aira et al. (2015) tras la determinación de la composición filogenética del estiércol de vaca y de cerdos, antes y después de su paso a través del intestino de Eisenia andrei difirió fuertemente en su composición, en la determinación de Proteobacterias esta disminuyó y aumentó en las deyecciones producidas por las lombrices tras ser alimentadas con estiércol de cerdo (antes: 60%, después: 32%) mientras que con el estiércol de vaca (antes: 24%, después: 82%). Mayor rendimiento en el crecimiento de las plantas se obtuvo en los vermicompost que recibieron inoculación microbiana (microorganismos benéficos y Bacillus sp.). Barea et al. (1976) indican que los microorganismos tienen la propiedad de producir hormonas vegetales y sustancias reguladoras del crecimiento de las plantas (PGR) que contribuyen a su desarrollo entre estas se citan a las auxinas, etileno, citoquininas, giberelinas y ácido abscísico que contribuyen en el desarrollo del cultivo (Barea et al., 1976). Además, de ello indica que se ha evidenciado que Bacillus sp., puede producir compuestos volátiles nematicidas tales como benceno acetaldehído, 2-nonanona, decanal, 2-undecanona y disulfuro de dimetilo, que controlan patógenos y nematodos permitiendo un mejor desarrollo de las plantas (Rao et al., 2017). Mahanta et al. (2012) obtuvieron mayor crecimiento de la planta, rendimiento de grano, contenido de clorofila de la hoja, al enriquecer vermicompost preparado a partir de pajilla de arroz con microorganismos promotores del crecimiento como son la Azotobacter chroococcum y Azospirillium brasilense. Al igual mayor altura registró Alvarez et al. (2018) al trabajar con consorcios microbianos obtenidos de raíces de plantas.
3.2.2. Peso de hipocótilo
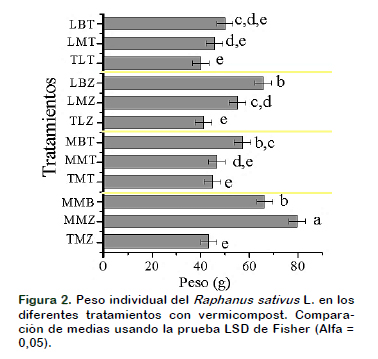
Los rabanitos cultivados no registraron presencia de plagas y ataque de enfermedades y obteniendo un alto rendimiento. El peso del hipocótilo en los diferentes tratamientos varió de 79,85 g en el tratamiento MMZ y 40,24 g en el tratamiento TLT (Figura 2). Amini et al. (2015) demostraron que la inoculación de Bacillus sp., y Pseudomonas sp., en el vermicompost aumentó el crecimiento y el rendimiento del azafrán, en condiciones de crecimiento orgánico. Cada tratamiento juega un papel directo en el crecimiento del hipocótilo como fuente de todos los macro-micronutrientes y sustancias húmicas. Para Gupta (2012) es posible que las diferencias en el crecimiento del fruto no este simplemente asociado a las diferencias en su contenido mineral nutricional, parece probable que haya otros insumos, como PGPR (Plant Growth-Promoting Rhizobacteria) que puede promover el crecimiento de raíces y brotes ya sea produciendo hormonas vegetales o metabolitos secundarios o aumentando la presencia de microorganismos beneficiosos o influyendo en sustancias que podrían estar involucradas en el crecimiento de las plantas y frutos.
4. Conclusiones
El manejo y gestión de residuos orgánicos peligrosos para la salud y contaminantes del medio ambiente, requieren de estrategias de desarrollo más rápidas para su revalorización. En base a los datos generados sobre las características físico químicas de los vermicompost obtenidos a partir de excreta de vacuno, se comprobó que es posible la obtención de un producto enriquecido y en menor tiempo (40 días) del habitual empleando el sistema zanja. El abono orgánico generado presentó efecto positivo en el desarrollo de la semilla, longitud del cultivo y peso del hipocótilo de Raphanus sativus L. En un futuro inmediato se recomienda estudiar el efecto que podría tener el enriquecimiento microbiano junto con Eisenia foetida a partir de otro tipo de excreta, amerita el empleo de la técnica de enriquecimiento microbiano debido a que su efecto combinado demostró tener un efecto significativo sobre la calidad del vermicompost. Sería útil reducir aún más el tiempo de descomposición previa explotando el potencial benéfico de los microorganismos generando un producto con mayor valor agregado.
Referencias bibliográficas
Aira, M.; Monroy, F.; Domínguez, J. 2007. Earthworms strongly modify microbial biomass and activity triggering enzymatic activities during vermicomposting independently of the application rates of pig slurry. Science of the Total Environment 385(1-3): 252-261. [ Links ]
Aira, M.; Bybee, S.; Pérez-Losada, M.; Domínguez, J. 2015. Feeding on microbiomes: Effects of detritivory on the taxonomic and phylogenetic bacterial composition of animal manures. FEMS Microbiology Ecology 91(11): 1-10. [ Links ]
Akbar Babael, A.; Goudarzi, G.; Neisi, A.; Ebrahimi, Z.; Alavi, N. 2016. Vermicomposting of cow dung, kitchen waste and sewage sludge with bagasse using Eisenia fetida. J Adv Environ Health R 4(42): 88-94. [ Links ]
Ali, T.; Mohammad, M.N.; Mohammad, P. 2018. Effects of vermicompost and vermiwash biofertilizers on fenugreek (Trigonella foenum) plant. Communications in Soil Science and Plant Analysis. USA.1-10 . [ Links ]
Alvarez, M.; Tucta, F.; Quispe, E.; Meza, V. 2018. Incidence of the inoculation of beneficial microorganisms in the strawberry (Fragaria sp.) crop. Scientia Agropecuaria 9(1): 33-42. [ Links ]
Amini, S.; Farahani Maleki, S.; Sharghi, Y.; Zahedi, H. 2015. Influence of vermicompost and bacterium of Bacillus and Pseudomonas on growth , yield and morphological traits of saffron . [ Links ]
Atiyeh, R.M.; Subler, S.; Edwards, C.A.; Bachman, G.; Metzger, J.D.; Shuster, W. 2000. Effects of vermicomposts and composts on plant growth in horticultural container media and soil. Pedobiologia 44(5): 579-590. [ Links ]
Balasubramani, R.; Sang, R.L.; Soon, W.C.; Dinh Duc, N.; Wood J.C.W.; Balamuralikrishnan, B.; Hupenyu Allan, M.; Mariadhas, V.A.; Naif, Abdullah, A-D; Ganesan, S. 2019. Positive e ff ects of compost and vermicompost produced from tannery wasteanimal fl eshing on the growth and yield of commercial crop-tomato (Lycopersicon esculentum L.) plant. Journal of Environmental Management 234: 154-158. [ Links ]
Barea, J.M.; Navarro, E.; Montoya, E. 1976. Production of plant growth regulators by Rhizosphere phosphate solubilizing bacteria. J of Applied Bacteriology 40(2): 129-134. [ Links ]
Barillari, J.; Cervellati, R; Costa, S; Guerra, MC; Speroni, E; Utan, A; Iori, R. 2006. Antioxidant and choleretic properties of Raphanus sativus L. sprout (Kaiware Daikon) extract. Journal of Agricultural and Food Chemistry 54(26): 9773-9778. [ Links ]
Bhat, S.A.; Singh, J.; Vig, A.P. 2016. Effect on growth of earthworm and chemical parameters during vermicomposting of pressmud sludge mixed with cattle dung mixture. Procedia Environmental Sciences 35: 425- 434. [ Links ]
Black, C.A.; Evans's, D.O.; Ensmunger, L.E.; White, J.L.; Clark, F.E.; Dineure, R.C. 1965. Methods of Soil Analysis II, Chemical and Microbiological Properties. American Soc. Argon. Madison, Wisconsin, USA. [ Links ]
Chidavaenzi, M.; Jere, M.; Bradley, M. 1997. Water and sanitation for all : partnerships and innovations Pit latrine effluent infiltration into groundwater. 23rd WEDC Conference, 1–5 September 1997: 59-62. [ Links ]
Das, D.; Bhattacharyya, P.; Ghosh, BC.; Banik, P. 2016. Bioconversion and biodynamics of Eisenia foetida in different organic wastes through microbially enriched vermiconversion technologies. Ecological Engineering 86: 154-161. [ Links ]
Diacono, M.; Montemurro, F. 2010. Long-term effects of organic amendments on soil fertility: A review. Agronomy for Sustainable Development 30: 401-422. [ Links ]
Edwards, C.; Scott, S.; Arancon, N. 2011. Quality criteria for vermicompost. Vermiculture technology. United States of America, CRC Press, p. 301. [ Links ]
Elvira, C.; Sampedro, L.; Benítez, E.; Nogales, R. 1998. Vermicomposting of sludges from paper mill and dairy industries with Eisena andrei: A pilot-scale study. Bioresource Technology 63(3): 205-211. [ Links ]
Etesami, H.; Emami, S.; Alikhani, H.A. 2017. Potassium solubilizing bacteria (KSB): mechanisms, promotion of plant growth, and future prospects • A review. Journal of soil science and plant nutrition 17(4): 897-911.
Fernández-Gómez, M.J.; Nogales, R.; Insam, H.; Romero, E.; Goberna, M. 2010. Continuousfeeding vermicomposting as a recycling management method to revalue tomato-fruit wastes from greenhouse crops. Waste Management 30(12): 2461-2468. [ Links ]
Ghosh, P.; Dash, P.K.; Sarker, R.; Mannan, A. 2014. Effect of salinity on germination, growth and yield of radish (Raphanus Sativus L.) varieties. International Journal of Biosciences (IJB) 5(1): 37-48. [ Links ]
Gupta, V. 2012. Beneficial Microorganisms for sustainable agriculture. 2012: 347-369.
Hernández, A.J.; Chacin, L.; Avila, J.; El khatib, N.; Chirinos, I.; Bracho, B. 2011. Methods salinity management of cattle dung for vermicomposting with the red worm ( Eisenia andrei ) Introducción. Revista Facultad de Agronomia 1: 342-350. [ Links ]
Higa, T.; Parr, J.F. 1989. Microorganismos benéficos y efectivos para una agricultura medio ambiente sostenible. 1989: 1-14.
Hughes, R.J.; Nair, J.; Ho, G. 2009. The risk of sodium toxicity from bed accumulation to key species in the vermifiltration wastewater treatment process. Bioresource technology 100(16): 3815-3819. [ Links ]
Karaca, A. 2011. Biology of earthworms. Turkey, Springer, Berlin, Heidelberg, v.24, 332 pp. [ Links ]
Karsten, G.; Drake, HL. 1995. Comparative assessment of the aerobic and anaerobic microfloras of earthworm guts and forest soils. Applied and Environmental Microbiology 44: 1039-1044. [ Links ]
Kaviraj; Sharma, S. 2003. Municipal solid waste management through vermicomposting employing exotic and local species of earthworms. Bioresource technology 90(2): 169-173. [ Links ]
Kostecka, J; Paczka, G. 2006. Possible use of earthworm Eisenia fetida (Saving.) biomass for breeding aquarium fish. European Journal of Soil Biology 42: 231-233. [ Links ]
Kumar, R.; Verma, D.; Singh, B.L.; Kumar, U.; Shweta. 2011. Composting of sugar-cane waste by-products through treatment with microorganisms and subsequent vermicomposting. Bioresource Technology 101(17): 6707-6711. [ Links ]
Lazcano, C; Gómez-Brandón, M; Domínguez, J. 2008. Comparison of the effectiveness of composting and vermicomposting for the biological stabilization of cattle manure. Chemosphere 72(7): 1013-1019. [ Links ]
Lee, J-K; Park, I; Choi, Y-J; Cho, J-S. 2012. Bacillus strains as feed additives: In vitro evaluation of its potential probiotic properties. Revista Colombiana de Ciencias Pecuarias 25: 577-585. [ Links ]
Lim, P.N.; Wu, T.Y.; Clarke, C.; Nik Daud, N.N. 2015. A potential bioconversion of empty fruit bunches into organic fertilizer using Eudrilus eugeniae. Int Journal of Environmental Science and Technology 12(8): 2533-2544. [ Links ]
Lim, S.L.; Lee, L.H.; Wu, T.Y. 2016. Sustainability of using composting and vermicomposting technologies for organic solid waste biotransformation: Recent overview, greenhouse gases emissions and economic analysis. Journal of Cleaner Production 111: 262-278. [ Links ]
Mahanta, K.; Jha, D.K.; Rajkhowa, D.J.; Kumar, M. 2012. Microbial enrichment of vermicompost prepared from different plant biomasses and their effect on rice (Oryza sativa L.) growth and soil fertility. Biological Agriculture & Horticulture 28(4): 241-250. [ Links ]
Mahmud, M.; Abdullah, R.; Jamilah Syafawati, Y. 2018. Effect of vermicompost amendment on nutritional status of sandy loam soil, growth performance, and yield of pineapple (Ananas comosus var. MD2) under field conditions. Malasia [ Links ]
Maji, D.; Misra, P.; Singh, S.; Kalra, A. 2017. Humic acid rich vermicompost promotes plant growth by improving microbial community structure of soil as well as root nodulation and mycorrhizal colonization in the roots of Pisum sativum. Applied Soil Ecology 110: 97-108. [ Links ]
Majlessi, M; Eslami, A; Najafi Saleh, H; Mirshafieean, S; Babaii, S. 2012. Vermicomposting of food waste: Assessing the stability and maturity. Iranian Journal of Environmental Health Science and Engineering 9(25): 1-6. [ Links ]
Mitchell, A. 1997. Production of Eisenia fetida and vermicompost from feed-lot cattle manure. Soil Biology and Biochemistry 29(3-4): 763-766. [ Links ]
Mousavi, S.A; Faraji, M.; Janjani, H. 2017. Recycling of three different types of rural wastes employing vermicomposting technology by Eisenia fetida at low temperature. 19(4): 601-606. [ Links ]
Nurhidayati, N.; Masyhuri, M; Indiyah, M. 2018. Direct and residual effect of various vermicompost on soil nutrient and nutrient uptake dynamics and productivity of four mustard Pak ‑ Coi (Brassica rapa L.) sequences in organic farming system. International journal of recycling of organic waste in agriculture. [ Links ]
Nagavallemma, K.P.; Wani, S.P.; Lacroix, S.; Rao M.B.; Sahrawat, K.L. 2006. Vermicomposting: Recycling Wastes into Valuable Organic Fertilizer. SAT eJournal ICRISAT 2(8): 1-14. [ Links ]
Natchimuthu, K; Periasamy, V; Mani, P; J. Arockia, JP. 2019. Vermicomposting of paper industry sludge with cowdung and green manure plants using Eisenia fetida: A viable option for cleaner and enriched vermicompost production. Journal of Cleaner Production. India [ Links ]
Ndegwa, P.M.; Thompson, S.A.; Das, K.C. 2000. Effects of stocking density and feeding rate on vermicomposting of biosolids. Bioresource Technology 71(1): 5-12. [ Links ]
OCDE-FAO (Food and Agriculture Organization of the United Nations). 2017. Carne: situación del mercado. 2017. [ Links ]
Olle, M. 2018. The effect of vermicompost on teh growth and quality of cress (Lepidium sativum). 25-28. [ Links ]
Patidar, A.; Gupta, R.; Tiwari, A. 2013. Potential of microbial inoculated water hyacinth amended thermophilic composting and vermicomposting in biodegradation of agro-industrial waste. Journal of Bioremediation & Biodegradation 4: 191. [ Links ]
Pinos-Rodríguez, J.M.; García-López, J.C.; PeñaAvelino, L.Y.; Rendón-Huerta, J.A.; GonzálezGonzález, C.; Tristán-Patiño, F. 2012. Impactos y regulaciones ambientales del estiércol generado por los sistemas ganaderos de algunos países de América. Agrociencia 46(4): 359-370. [ Links ]
Rao, M.S.; Kamalnath, M.; Umamaheswari, R.; Rajinikanth, R.; Prabu, P.; Priti, K.; Grace, G.N.; Chaya, M.K.; Gopalakrishnan, C. 2017. Bacillus subtilis IIHR BS-2 enriched vermicompost controls root knot nematode and soft rot disease complex in carrot. Scientia Horticulturae 218: 56-62. [ Links ]
Saravana, S.; Aruna, D. 2013. Nutrient enrichment of vermicompost by probiotics supplementation. European Journal of Experimental Biology 3(4): 84-88. [ Links ]
Schuldt, M. 2006. Lombricultura. Teoría y práctica. Ed. M Prensa. Madrid, s.e., 306 pp. [ Links ]
Schuldt, M; Rumi, A; Gutiérrez, DE. 2005. Determinacion de ¨edades¨ (clases) en poblaciones de Eisenia foetida (Annelida : Lumbricidae) y sus implicancias reprobiológicas. Revista del Museo de La Plata 17: 1-10. [ Links ]
Sierra, J.; Desfontaines, L.; Faverial, J.; Loranger-Merciris, G.; Boval, M. 2013. Composting and vermicomposting of cattle manure and green wastes under tropical conditions: Carbon and nutrient balances and endproduct quality. Soil Research 51(2): 142- 151. [ Links ]
Sifolo, S.C.; Flavien, E.; Kouadio, I.K.; Barsan, N.; Nedeff, V.; Bi, Z. 2018. Vermicompost utilization : A way to food security in rural area. Heliyon, Romania. [ Links ]
Sharma, A.; Saha, T.; Arora, A.; Shah, R.; Nain, L. 2017. Efficient Microorganism compost benefits plant growth and improves soil health in calendula and marigold. Horticultural Plant Journal 3(2): 67-72. [ Links ]
Singh, A; Sharma, S. 2002. Composting of a crop residue through treatment with microorganisms and subsequent vermicomposting. Bioresource technology 85(2): 107-111. [ Links ]
Sultana, S.; Kashem, M.A.; Mollah, A.K.M. 2015. Comparative assessment of cow manure vermicompost and npk fertilizers and on the growth and production of Zinnia (Zinnia elegans) Flower. Open Journal of Soil Science 05(09): 193-198. [ Links ]
Sumi, C.D.; Yang, B.W.; Yeo, I-C; Hahm, Y.T. 2015. Antimicrobial peptides of the genus Bacillus : a new era for antibiotics. Canadian Journal of Microbiology 61(2): 93-103. [ Links ]
Suthar, S. 2009. Vermicomposting of vegetablemarket solid waste using Eisenia fetida: Impact of bulking material on earthworm growth and decomposition rate. Ecological engineering 35(5): 914-920. [ Links ]
Suthar, S; Singh, S. 2008a. Comparison of some novel polyculture and traditional monoculture vermicomposting reactors to decompose organic wastes. Ecological engineering 33(34): 210-219. [ Links ]
Suthar, S; Singh, S. 2008b. Feasibility of vermicomposting in biostabilization of sludge from a distillery industry. Science of the total environment 394(2-3): 237-243. [ Links ]
Tripathi, G; Bhardwaj, P. 2004. Comparative studies on biomass production, life cycles and composting efficiency of Eisenia fetida (Savigny) and Lampito mauritii (Kinberg). Bioresource Technology 92(3): 275-283. [ Links ]
Vázquez, J.; Loli, O. 2018. Compost and vermicompost as amendments in the recovery of a soil degraded by the management of Gypsphila paniculata. Scientia Agropecuaria 9(1): 43-52. [ Links ]
Wang, K.; He, C.; You, S.; Liu, W.; Wang, W.; Zhang, R.; Qi, H.; Ren, N. 2015. Transformation of organic matters in animal wastes during composting. Journal of hazardous materials 300: 745-753 . [ Links ]
Wang, S.; Fu, B.J.; Gao, G.Y.; Yao, X.L.; Zhou, J. 2012. Soil moisture and evapotranspiration of different land cover types in the Loess Plateau, China. Hydrology and Earth System Sciences 16(8): 2883-2892. [ Links ]
Wani, K.A.; Mamta; Rao, R.J. 2013. Bioconversion of garden waste, kitchen waste and cow dung into value-added products using earthworm Eisenia fetida. Saudi Journal of Biological Sciences 20(2): 149-154. [ Links ]
Corresponding author
E-mail: svelecela@ucacue.edu.ec (S. Velecela)
Received November 3, 2018.
Accepted April 28, 2019.

