











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El camarón Cryphiops caementarius es la especie que se distribuye solo en los ríos costeros de Perú y Chile (Moscoso, 2012). En Chile, esta especie es considerada como vulnerable e incluso en peligro de extinción en la región Metropolitana (Bahamonde et al., 1998). En Perú, la especie soporta extracción comercial, dada su abundancia poblacional principalmente en los ríos de Arequipa, desde donde se extrajo alrededor de mil toneladas, para abastecer principalmente al mercado de Lima (Wasiw y Yépez, 2015; Produce, 2018). En ambos países es considerada como una especie de preocupación (De Grave et al., 2013) y por ello hay interés por el cultivo comercial, habiéndose realizado investigaciones en producción de postlarvas (Meruane et al., 2006; Romero et al., 2013), en cultivo de adultos en sistemas individuales (Reyes, 2016) y en sistema de biofloc (Ulloa et al., 2019), así como en aspectos ecológicos (Escobar et al., 2017; Reyes et al., 2020) y nutricionales (Acosta et al., 2018; Terrones y Reyes, 2018; Ramírez et al., 2018), entre otros.
El camarón C. caementarius, es omnívoro no selectivo, que se alimenta de restos de vegetales, microalgas (Clorofitas, Cianofitas y Diatomeas), restos de moluscos, larvas de insectos (Efemerópteros, Dípteros, Quironómidos), copépodos y otros crustáceos (Bahamonde y Vila, 1971). Los camarones adultos aceptan alimento balanceado cuyos insumos son harina de pescado y harinas de soya, de trigo, de maíz, entre otros (Reyes, 2016; Acosta et al., 2018).
En el cultivo de camarones, el alimento constituye entre 40% y 60% del costo opera tivo de la producción (Sarman et al., 2018), donde la harina de pescado es la principal fuente de proteínas, pero encarece el costo de producción. Los carbohidratos son la fuente de energía primaria en las dietas de animales y es la menos costosa en las dietas de animales acuáticos (Wang et al., 2016). Los crustáceos de agua dulce como Macrobrachium rosenbergii y M. nipponense, son capaces de digerir los carbohidratos complejos (Wang et al., 2016) como el almidón de maíz crudo y pregelatinizado (Kong et al., 2019). El maíz amarillo (Zea mays), contiene hasta 73% de almidón y las proteínas oscilan entre 7% y 11% que se encuentra en el endospermo (Gwirtz y García-Casal, 2014).
En C. caementarius se ha formulado una dieta para engorde (Reyes, 2016), donde el uso del 15% de ensilado biológico de residuos blandos de molusco en reemplazo de la harina de pescado, no afecta el crecimiento (Terrones y Reyes, 2018). El ensilado biológico mejora la digestibilidad, el contenido nutricional y la palatabilidad de las dietas (Coelho-Emerenciano y Massamitu-Furuya, 2006), dado a las bacterias lácticas como Lactobacillus, Streptococcus, Pediococcus, entre otras (Muck et al., 2018), que al producir sustancias antimicrobianas reducen las bacterias patógenas (Hoseinifar et al., 2018). Estas bacterias lácticas contribuyen con el proceso digestivo, la asimilación y con el crecimiento, como en postlarvas de M. inca (Dávila et al., 2013).
El ensilado biológico de harina de maíz (24%) en la dieta de M. rosenbergii reem plaza a la harina de maíz, sin afectar el crecimiento en peso (Coelho-Emerenciano y Massamitu-Furuya, 2006). En C. caemen-tarius, no se conoce si este tipo de ensilado mejora el crecimiento. Por consiguiente, el objetivo fue evaluar dietas con ensilado biológico de harina de maíz en el mejoramiento del desempeño productivo del camarón de río C. caementarius.
Materiales y métodos
Los camarones se capturaron del río Pativilca cerca del Centro Poblado Huayto (10°39''50'' S, 77°39'22'' O) (Lima, Perú) y se transportaron en un sistema individual donde cada camarón se introdujo en un vaso de plástico de 200 mL (los vasos se agujeraron para el flujo de agua) los que se acondicionaron en cajas de plástico (45 L) con agua del mismo río y con aireación (Reyes, 2016). La densidad de transporte fue de 60 camarones por caja. El transporte terrestre duró 4 h y no hubo mortalidad. Los camarones se aclimataron, en laboratorio, por 10 días en el mismo sistema de transporte y se alimentaron con la dieta basal (28% proteína cruda). Los cambios de agua (30%) y la extracción de restos de alimento y de desechos sólidos de excreción se realizaron cada tres días con sifón.
La especie C. caementarius se reconoció según Méndez (1981) y el sexo de los machos se verificaron por la presencia de gonóporos en los coxopoditos del quinto par de periópodos, además se diferenciaron por el tamaño de las quelas y la amplitud del abdomen (Reyes et al., 2018).
El sistema de cultivo individual fue el empleado por Reyes (2016), el cual consistió de 12 acuarios de vidrio (0,60 m de largo; 0,31 m de ancho; 0.35 m de alto; de 0,186 m2 y 55 L). En cada acuario el sistema de recirculación de agua fue del tipo air-water-lift (1,5 L min-1) con filtro biológico percolador (2,5 L) con capas de esponja sintética, grava y conchuelas trituradas, además de difusores para circulación y aireación del agua. Dentro de cada acuario, se instalaron seis recipientes de plástico (19 cm de diámetro; 8 cm de altura y de 284 cm2) que se dispusieron en dos grupos de tres niveles. Los recipientes tuvieron orificios en los laterales para circulación de agua y en un extremo se instaló un tubo vertical (PVC de ½'' Ø) por donde se introdujo los gránulos de alimento. En cada recipiente de cultivo se sembró un camarón que equivalió a seis camarones por acuario (32 camarones m-2) y por ello, fueron empleados 72 camarones machos (4,90 cm a 5,03 cm de longitud total) que mostraron apéndices cefalotorácicos completos.
El ensilado se elaboró con el grano seco de maíz (Z. mays) el cual se molió, tamizó (120 μm) y fue mezclado con 10% de melaza, 15% de inóculo de bacterias lácticas y 70% de agua hervida tibia (50ºC). El homogenizado se introdujo en frascos de vidrio (1 L) y fue incubado a 40°C por 48 h hasta pH 4,4. El ensilado fue secado a 60ºC por 24 h, molido y tamizado (120 μm). El maíz amarillo duro se adquirió del mercado de abastos La Perla (Chimbote, Perú). La melaza de caña de azú car (75° Brix) se obtuvo de la Empresa Agroindustrial San Jacinto SAC (Nepeña, Ancash, Perú). Las bacterias lácticas comerciales liofilizadas Lyofast Y 456 B-1UC (Lactobacillus delbruecki y Streptococcus thermophilus) se activaron según Berenz (1996).
La dieta basal fue la formulada por Reyes (2016). En las dietas experimentales se reemplazó la harina de maíz por el ensilado de harina de maíz (Tabla 1). Las dietas se peletizaron a temperatura ambiente en una prensa comercial. Los pellets se secaron a 50ºC por 24 h, luego se quebraron (3 mm) y almacenaron en bolsas de plástico de cierre hermético. La hidroestabilidad de los gránulos de las dietas se determinó según Zettl et al. (2019). La tasa de alimentación de los camarones fue de 6% del peso húmedo por día y se reajustó después de cada muestreo. El alimento fue proporcionado a las 08:00 h (40%) y 18:00 h (60%).
El análisis químico proximal del ensilado y de las dietas comprendió proteínas, lípidos y fibra que se realizó en el Laboratorio Certificado COLECBI el cual empleó el método UNE-EN ISO 5983-2 Parte 2 dic.2006 con factor de 6,25 para el contenido proteico. Además, la humedad, lípidos, cenizas y fibra se determinó por el método de la A.O.A.C. (1990). Para los carbohidratos se empleó la fórmula por diferencia: 100 - (% proteínas + % grasas + % fibra + % ceniza). El pH del ensilado de maíz se registró con pH metro di gital (± 0,01) y según el método de Betancourt et al. (2014).
Los camarones fueron muestreados cada 30 días. El peso total se determinó en balanza digital ADAM AQT 600 (± 0,01 g). La longitud total se midió con regla graduada (± 0,5 mm) y se consideró desde la escotadura postorbital hasta el extremo posterior del telson. El desempeño productivo se determinó con los siguientes parámetros:
Ganancia Porcentual = (X2 - X1/X1)*100
Tasa de Crecimiento Absoluto = X2 - X1/t2 - t1
Tasa de Crecimiento Específico = [ln X2 - ln X1) / t2 - t1] * 100 (g)
Supervivencia = (Ni*100)/No
Donde, X1 y X2 fue el peso húmedo (g) o la longitud total (cm), inicial y final; t1 y t2 la du ración en días; ln X1 y ln X2 fue el logaritmo natural del peso o la longitud inicial y final. No y Ni es el número inicial y final de camarones sembrados, respectivamente.
Los desechos sólidos acumulados en los acuarios se extrajeron con sifón dos veces por semana. La calidad del agua se monitoreó semanalmente y se determinó oxígeno disuelto y temperatura con Oxímetro digital Sension8 (± 0,01 mg L-1; ± 0,01 ºC), el pH con pH-metro digital 110 (± 0,01 unidades), y el amonio total y los nitritos se determinaron semanalmente con kit Sera Aqua-test box (± 0,1 mg L-1).
Los datos se analizaron con la prueba de Kolmogorov-Smirnov y todos cumplieron con el supuesto de distribución normal. Las diferencias entre las medias de los tratamientos se determinaron por análisis de varianza de una vía y con la prueba de Duncan.
Tabla 1 Composición porcentual de dietas para C. caementarius, de acuerdo a la proporción de ensilado biológico de harina de maíz
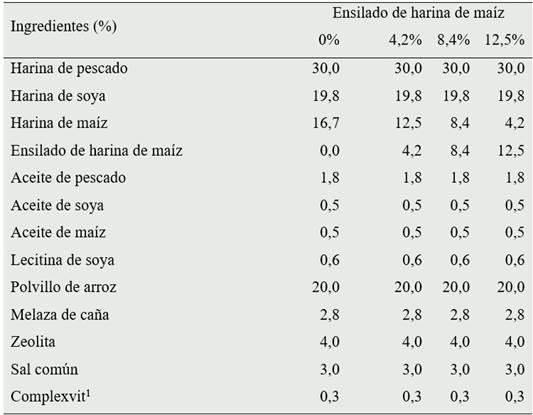
1 Comprende (kg-1): Vitaminas A 8 g; E 7 g; B1 8 g; B2 16 g; B6 11,6 g; B12 0,02 g; C 5 g; D3 5 g; K3 1 g; Nicotinamida 10 g; Niacina 6 g; Biotina 0,3 g; DL Metionina 20 g; Pantotenato de calcio 47 g; Cloruro de sodio 2.7 g; Cloruro de potasio 34 g; Sulfato de magnesio 7 g; Maca 5 g: y Excipientes 1 g.
Tabla 2 Calidad del agua de cultivo de C. caementarius alimentados con dietas con diferentes proporciones de ensilado biológico de harina de maíz, durante 60 días
| Ensilado de harina de maíz | Temperatura (ºC) | Oxígeno (mg L-1) | pH | Amonio total (mg L-1) | Nitritos (mg L-1) |
| 0% 4,2% 8,4% 12,5% | 25,50 ± 0,43a 25,50 ± 0,25a 25,75 ± 0,00a 25,58 ± 0,38a | 6,38 ± 0,10a 6,29 ± 0,03a 6,37 ± 0,06a 6,32 ± 0,02a | 8,12 ± 0,08a 8,14 ± 0,09a 8,11 ± 0,04a 8,13 ± 0,03a | 0,02 ± 0,01a 0,03 ± 0,01a 0,02 ± 0,01a 0,03 ± 0,01a | 0,02 ± 0,00b 0,03 ± 0,01ab 0,03 ± 0,00a 0,03 ± 0,01a |
Datos con letras diferentes en superíndices en una misma columna indica diferencia significativa (p < 0,05).
El nivel de significancia fue del 5%. El procesamiento estadístico se realizó con el programa estadístico SPSS versión 23 para Windows.
Resultados y discusión
Los parámetros de calidad del agua de cultivo del camarón C. caementarius fueron similares (p > 0,05) entre tratamientos, excepto los nitritos que fue significativamente (p < 0,05) mayor en el agua donde los camarones se alimentaron con dietas con ensilado (Tabla 2). Sin embargo, la calidad del agua de los acuarios, estuvo dentro del rango informado para el ambiente donde habita la especie de camarón (Wasiw y Yépez, 2015).
El ensilado de harina de maíz (Tabla 3) tuvo bajo contenido de proteínas (8,57%), aunque los lípidos totales (16,49%) y las cenizas (2,25%) se mantuvieron en niveles cercanos al de los gránulos de maíz (Gwirtz y García-Casal, 2014). En cambio, los carbohidratos del ensilado (71,24%) fueron menores a la harina de maíz que contiene entre 84% a 90% de carbohidratos digeribles (Zainuddin et al., 2014). Esto sugiere que durante el proceso de fermentación láctica se utilizaron parte de los carbohidratos del maíz y es más probable que hayan sido los de la melaza que posee sacarosa (68,36%) como azúcar principal, seguida de glucosa (18,50%) y maltosa (13,14%) (El-Gendy et al., 2013), que son utilizados como sustratos para el crecimiento de Lactobacillus (Barrazueta et al., 2019). Esto explicaría el mayor contenido de carbohidratos de las dietas conforme se incrementó la proporción del ensilado.
El incremento de carbohidratos en las dietas experimentales permitió dar mayor hi droestabilidad a los gránulos de alimento (Tabla 3), lo que debe haber evitado la lixi viación de los nutrientes. De acuerdo con Xiao et al. (2014), el mayor nivel de almidón de maíz en las dietas de Procambarus clarkii, ocasiona que estas sean más estables en el agua debido al efecto de unión del almidón.
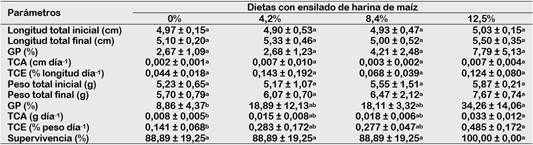
El crecimiento en longitud de los camarones fue similar (p > 0,05) entre tratamientos (Figura 1A, Tabla 4). De igual manera, sucedió con el crecimiento en peso, aunque hubo tendencia a incrementar a partir de los 30 días de cultivo, con las mayores proporciones de ensilado (Figura 1B). Es probable que se requiera mayor tiempo de cultivo para observar resultados diferentes en el peso de los camarones entre tratamientos. En cambio, los parámetros de crecimiento en peso (GP, TCA y TCE) fueron significativamente mayores (p < 0,05) con 12,5% de ensilado en relación con la dieta control, pero fue similar (p > 0,05) con 4,2% y 8,4% de ensilado (Tabla 4). Estos resultados, demuestran por primera vez, que el crecimiento en peso de C. caementarius fue mayor cuando se alimentaron con dietas que tuvieron ensilado de harina de maíz, en comparación con los alimentados sin ensilado. Además, la harina de maíz (16,7%) de la dieta puede ser reemplazada hasta 75% de ensilado, es decir con 12,5% de ensilado de dicho insumo, sin afectar el crecimiento del camarón adulto. Los resultados de la investigación son consistentes con los obtenidos en postlarvas de M. rosenbergii cuyo crecimiento en peso fue mayor con el reemplazo total de la harina de maíz por el de ensilado de harina de maíz (24%) en la dieta, y la que tuvo una mayor palatabilidad (Coelho-Emerenciano y Massamitu-Furuya, 2006).
Los parámetros de crecimiento en peso de C. caementarius fueron mayores, conforme se incrementó el ensilado de harina de maíz (4,2% a 12,5%) en la dieta, aunque sin diferencias (p > 0,05) entre ellos, pero si con el control. Estos resultados indicarían que este ensilado debe contener algunas sustancias que contribuyeron con la digestión o asimilación de nutrientes por el camarón. El ensilado de maíz posee aminas biogénicas como cadaverina, putrescina, espemina y espermidina, debido a la acción de las bacterias ácido lácticas durante el proceso fermentativo (Barbieri et al., 2019). Aunque, no fue determinado estas aminas en las dietas experimentales, sin embargo, se conoce que la espermina es promotora del crecimiento en Litopenaeus stylirostris (Tapia-Salazar et al., 2001).
Tabla 3 Composición química proximal y características físicas y organolépticas de dietas con diferentes proporciones de ensilado biológico de harina de maíz
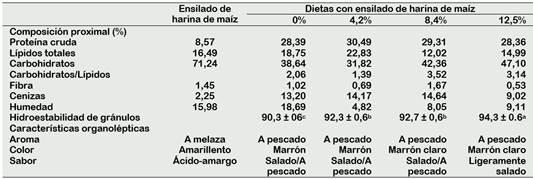
Datos con letras diferentes en superíndice en una misma fila indica diferencia significativa (p < 0,05).
Las bacterias del ensilado (L. delbrueckii y S. thermophilus), por el hecho de haber sido incubadas a 40ºC y procesadas a 50ºC, deben haber permanecido vivas, que al ser ingeridas por el camarón C. caementarius podrían haber contribuido con mejorar la flora microbiana y con ello la asimilación de nutrientes, como lo sugerido para la misma especie cuando se alimentó con dietas con ensilado procesado a 40 ºC (Terrones y Reyes, 2018). La temperatura óptima para el crecimiento y producción de ácido láctico por Lactobacillus delbrueck subsp. bulgaricus es de 44 ºC (Aghababaie et al., 2014). En esta condición, Lactobacillus por ser homofermentativo, tiene una velocidad de fermentación más rápida, una proteólisis reducida, mayores concentraciones de ácido láctico, menores contenidos de ácido acético y butírico, menor contenido de etanol y mayor recuperación de energía y materia seca (Santos et al., 2013). El ácido láctico, obtenido de la fermentación del almidón de maíz por especies de Lactobacillus, inhibe el crecimiento de levaduras y bacterias oportunistas con lo cual se mejora la estabilidad aeróbica del ensilado (Kleinschmit y Kung, 2006) y contribuye con la flora benéfica del huésped.
La harina de maíz, en las dietas experimentales, solo fue reemplazada por el ensilado de harina de maíz, sin modificar la proporción de los otros insumos, lo que permitió mantener mínima variación de proteínas de las dietas (28% a 30%). De acuerdo con Arcari et al. (2016), el contenido de proteína bruta del maíz molido húmedo no se altera durante el tiempo de ensilado; es por ello que el contenido de proteínas (8,57%) del ensilado de maíz, estuvo dentro del rango (7% a 11%) del contenido de maíz amarillo (Gwirtz y García-Casal, 2014), lo que confirmaría que no hubo proteólisis durante el ensilado o esta fue muy reducida. Las especies de Lactobacillus ocasionan reducida proteólisis durante el ensilado de maíz (Santos et al., 2013). Además, por el hecho de que durante el proceso del ensilado hubo acidez del medio (pH 4,40) generada por las bacterias ácido lácticas, sugiere que la melaza o los carbohidratos de las otras harinas empleadas en la dieta, hayan contribuido con la fermentación hasta disminuir el pH del medio.
Las bacterias y las enzimas del núcleo del maíz, contribuyen con la proteólisis de la matriz y de la superficie del gránulo del almidón del maíz, así como a su digestibilidad (Junges et al., 2017), lo cual podría estar limitada en la harina de maíz empleada, toda vez que se conoce que las proteínas de la superficie de los gránulos de almidón, afectan la digestibilidad del almidón (Svihus et al., 2005). Estas investigaciones explicarían el menor crecimiento en peso de C. caementarius alimentado con la dieta control (sin ensilado), que aunado a la baja proporción de harina de maíz (16,7%) empleada, ocasionó bajo contenido de carbohidratos (38,64%) en dicha dieta. En M. idae el mayor crecimiento se logra con dietas que contienen entre 29% y 37% de harina de maíz, cuya digestibilidad fue de 76,9% la cual es usada como fuente de energía (Sundaravadivel y Sethuramalingam, 2017). Es conveniente investigar en el camarón C. caementarius, la digestibilidad aparente de la harina de maíz y del ensilado biológico de la harina de maíz.
En las dietas experimentales, el incremento del ensilado de harina de maíz ocasionó aumento de carbohidratos y disminución de lípidos, y la proporción carbohidratos/lípidos incrementó de 1,39 a 3,14 con la proporción de ensilado (4,2% y 12,5%, respectivamente) (Tabla 3). Además, las mayores tasas de crecimiento en peso de C. caementarius se obtuvo con 12,5% de ensilado en la dieta, cuya proporción de carbohidratos/lípidos fue de 3,14:1, que correspondió a 47,10% de carbohidratos y de 14,99% de lípidos.
Tabla 4 Parámetros de crecimiento en longitud y peso de C. caementarius alimentado con dietas con diferentes proporciones de ensilado biológico de harina de maíz, durante 60 días
En Cherax quadricarinatus mayor crecimiento se logra con dietas cuya proporción de carbohidratos/lípidos fue de 3,60:1, que corresponde a 29% de carbohidratos y 8% de lípidos (Zhu et al., 2013). En consecuencia, los resultados sugieren que los machos adultos de C. caementarius utilizan los carbohidratos como fuente de energía en vez de las proteínas en la dieta, lo que explica los mayores parámetros de crecimiento en peso del camarón.
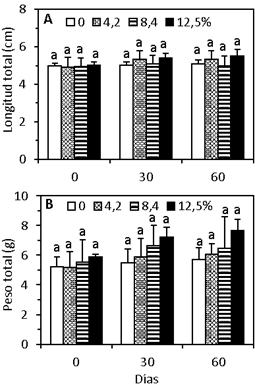
Figura 1 Crecimiento en longitud (A) y peso (B) de C. caementarius alimentados con diferentes proporciones de ensilado biológico de harina de maíz en la dieta, durante 60 días.
Salgado-Leu y Tacon (2015), determinaron en juveniles de Samastacus spinifrons, que la dieta con 30% de proteínas y bajo contenido de carbohidratos (16,3% a 23,5%) ocasiona altos índices de crecimiento y una eficiente utilización de las proteínas de la dieta, lo que sugiere que la especie usa los carbohidratos dietarios como principal fuente de energía para ahorrar proteínas y maximizar el crecimiento. Los camarones machos adultos de C. caementarius tendrían capacidad de utilizar alto contenido de carbohidratos (47,10%) en la dieta que tuvo un nivel de proteínas de 28,36% (Tabla 3), y que, de acuerdo a los resultados de los parámetros de crecimiento en peso, indicaría que los carbohidratos de las dietas con ensilado estarían siendo utilizados como fuente de energía, dejando a las proteínas para la conformación de masa muscular.
De acuerdo con Xiao et al. (2014), cuando los niveles de carbohidratos en la dieta exceden el óptimo, los animales no crecen porque dicho exceso disminuye la capacidad de digerir, absorber y asimilar los carbohidratos de la dieta, incluso llegan a reducir la ingesta de alimento. Esto no sucedió en la investigación, aun cuando los carbohidratos de la dieta con alta proporción de ensilado (12,5%) fue de 47,10%, por lo que se conjetura que los machos adultos de C. caementarius requieren una dieta con alto contenido de carbohidratos sin afectar la ingesta del alimento ni el crecimiento, como ha sido informado en otras especies de crustáceos (Tabla 5), donde M. rosenbergii requiere 24% de ensilado de maíz (Coelho-Emerenciano y Massamitu-Furuya, 2006), P. clarkii 20% (Xiao et al., 2014) y M. nipponense 18% (Kong et al. 2019). Aunque no se conocen las enzimas digestivas de C. caementarius que digieran carbohidratos, sin embargo, el hecho de lograr mayores parámetros de crecimiento en peso con la dieta que contiene 12,5% de ensilado, sugiere que esta especie de camarón aprovecha los carbohidratos de dicha dieta; y es probable que fue estimulado por las bacterias del ensilado.
Tabla 5 Comparación de las mejores proporciones de ensilado biológico de maíz y de almidón de maíz en dietas, para crecimiento de crustáceos de agua dulce
| Tipo de carbohidrato | Proporción en dieta | Especie | Estado | Tamaño | Ref. |
| Ensilado de harina de maíz | 12,5% | C. caementarius | Adulto | 5,23 - 5,87 g | EE |
| Ensilado de harina de maíz | 24,0% | M. rosenbergii | Postlarva | 0,12 g | 1 |
| Almidón de maíz | 15% | M. nipponense | Postlarva | 0,13 g | 2 |
| Almidón de maíz crudo | 18% | M. nipponense | Postlarva | 0,12 g | 3 |
| Almidón de maíz pregelatinizado | 18% | M. nipponense | Postlarva | 0,12 g | 3 |
| Almidón de maíz | 20% | P. clarkii | Juveniles | 0,39 g | 4 |
EE. Este estudio. 1) Coelho-Emerenciano y Massamitu-Furuya, 2006. 2) Ding et al., 2016. 3) Kong et al. 2019. 4) Xiao et al., 2014.
En C. cainii, las mayores cantidades de microvellosidades por pliegue intestinal se deben a las bacterias del fermentado con L. casei que actúan como probióticos y son responsables de una mejor absorción y utilización de nutrientes (Siddik et al., 2020).
El almidón de maíz contiene entre 20% a 25% de amilosa y de 75% a 80% de amilopectina (Hamaker et al., 2019), donde la amilopectina es digerida relativamente bien por los camarones peneidos (Gaxiola et al., 2006) y es probable que sea similar en los carídeos. Se conoce que durante la hidrólisis del almidón se produce al azar ruptura de los enlaces α1-4 y α1-6, cuyos productos de degradación de la amilosa son la maltosa y maltotriosa, y de la amilopectina son la dextrina y oligómeros formados por enlaces α1-6 (Alcázar-Alay y Meireles, 2015). La amilopectina tiene un área de superficie, por molécula, mucho mayor que la amilosa, que lo convierte en un sustrato preferible para el ataque amilolítico (Singh et al., 2010). Estas investigaciones probablemente expliquen la utilización de los carbohidratos de las dietas por los camarones machos adultos de C. caementarius, toda vez que la especie es omnívora que, en su dieta natural, se alimen ta de microalgas y de restos de diversos vegetales y también de animales (Bahamonde y Vila, 1971), por lo que deben estar adaptados para metabolizar carbohi dratos. En postlarvas de M. nipponense las dietas con 18% de almidón de maíz crudo o pregelatinizado mejoran el crecimiento (Ding et al., 2016; Kong et al., 2019) y en P. clarkii con 20% de almidón de maíz (Xiao et al., 2014). Sin embargo, es conveniente continuar las investigaciones para entender mejor la nutrición de la especie con diferentes fuentes de carbohidratos en la dieta, principalmente para utilizarlos en el engorde del camarón.
Conclusiones
Los camarones machos adultos de C. caementarius utilizan dietas con alto contenido de ensilado de harina de maíz (12,5%), cuya proporción carbohidratos/lípidos fue de 3,14:1 lo que ocasionó mayores tasas de crecimiento en peso (p < 0,05) en relación con aquellos alimentados con dieta sin ensilado. La supervivencia fue alta (> 88%) e independiente de los tratamientos. Es conveniente continuar con las investigaciones con el uso del ensilado biológico de otras fuentes de carbohidratos en la dieta que permita mejorar el engorde del camarón.

