











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El término “Rizobacteria Promotora de Crecimiento Vegetal” (RPCV) ha sido usado para describir bacterias que se desarrollan en las raíces de las plantas y que provocan efectos positivos para las mismas (Kalita et al., 2015). Tales beneficios han sido aplicados en diferentes áreas como los biofertilizantes, control de enfermedades, biorremediación y biopesticidas (Adesemoye et al., 2008). Estos microorganismos pueden también reducir los efectos negativos causados por estrés en las plantas y contribuir a la rizorremediación (Porcel et al., 2014). Los mecanismos de acción de las RPCV pueden también subdividirse en mecanismos indirectos y directos (Shameer y Prasad, 2018). Los indirectos abarcan la resistencia sistémica adquirida y resistencia sistémica inducida, inhibición de agentes fitopatógenos (Singh et al., 2017). Los mecanismos directos comprenden la fijación de nitrógeno, inhibición de la síntesis de etileno, síntesis de hormonas vegetales, la solubilización de nutrientes como fósforo, hierro y potasio (Olanrewaju et al., 2017).
Los géneros predominantes que actúan como RPCV son Pseudomonas y Bacillus (Grover et al., 2011), las especies del género Bacillus son formadoras de esporas tolerantes al estrés, secretan exopolisacáridos y sideróforos que inhiben el movimiento de iones tóxicos y promueven el movimiento del agua en los tejidos vegetales lo que favorece al desarrollo de la planta (Radhakrishnan et al., 2017).
En la actualidad, el tomate riñón (Lycopersicum esculentum Mill.) es uno de los vegetal más cultivados y consumidos alrededor del mundo, alcanzando una producción de 182.258 millones de kg por año (ESPAC, 2018).
La gran demanda de tomate riñón y la exigencia del mercado han hecho que los productores se interesen por nuevos sistemas de producción que generen mayor rendimiento y productos de calidad (Espinosa et al., 2017), en este sentido la aplicación de RPCV en plántulas representa una alternativa para mejorar el desarrollo, rendimiento y la obtención de biomasa vegetal (Chaudhary et al., 2012; Martínez, 2011). En varios estudios se ha demostrado que la inoculación con RPCV ha logrado mejorar el crecimiento y desarrollo del cultivo de tomate riñón (Cabra et al., 2017; Martínez et al., 2013; Santillana et al., 2005; Terry y Galán, 2006).
Las RPCV que permite disminuir o sustituir la fertilización química, con la finalidad de mejorar las características de rendimiento y calidad, evitando la contaminación de recursos naturales dentro de los sistemas productivos agrícolas (Espinosa-Palomeque y Cano-Rios, 2019).
En este contexto, esta investigación tuvo como objetivo determinar la capacidad promotora de crecimiento de dos cepas de Bacillus en plántulas de tomate riñón (Lycopersicum esculentum Mill.) y de esta manera obtener una herramienta biotecnológica para el uso en la agricultura.
Materiales y métodos
La evaluación se llevó a cabo en Ecuador, provincia de Tungurahua, cantón Ambato, la cual se encuentra a 2590 m.s.n.m., una humedad media de 68,57% y 14,6 °C como temperatura media.
Las cepas utilizadas estaban identificadas previamente como pertenecientes al género Bacillus y fueron las siguientes: IB10, CT11, ambas cepas pertenecen al cepario del CIVABI, ubicado en la Universidad Politécnica Salesiana de la ciudad de Quito. Estas cepas en ensayos in vitro obtuvieron los mejores resultados en ensayos de solubilización de fósforo, producción de auxinas y fijación biológica de nitrógeno, resultados reportados en el trabajo de investigación de Mamarandi y Ojeda (2019).
Identificación molecular de las cepas de Bacillus
Los microorganismos bacterianos se cultivaron en Tripticasa Soya Agar (TSB) y se incubaron a 30 °C y 100 rpm. El ADN se extrajo a partir de cultivos de 24 horas mediante la metodología descrita por Sambrook y Russell (2001).
Para la identificación molecular de las especies, se utilizaron cebadores de genes específicos que sirven para identificar especies bacterianas del género Bacillus, detallados en Tabla 1 los cuales fueron descritos por Madslien et al. (2013) y Nayak et al. (2013).
La reacción de la cadena de polimerasa se realizó con el siguiente protocolo: desnaturalización inicial a 95 °C por 2 minutos, 24 ciclos de desnaturalización a 95 °C por 30 segundos, hibridación del cebador a 58 °C por 1 minuto, extensión inicial por 2 minutos a 72 °C y extensión final de 9 minutos a 72 °C, la muestra amplificada se mantuvo a 4 °C.
Tabla 1 Cebadores utilizados en la identificación molecular
| Región para amplificar | Secuencia 5´-3´ | Temperatura de hibridación (°C) |
| Bacillus licheniformis | ||
| Gen (lchAA) Cebador Forward | ACTGAAGCGATTCGCAAGTT | 58 |
| Gen (lchAA) Cebador Reverse | TCGCTTCATATTGTGCGTTC | |
| Bacillus megaterium | ||
| Gen (phaC) Cebador Forward | CGTGCAAGAGTGGGAAAAAT | 58 |
| Gen (phaC) Cebador Reverse | TCGCAATATGATCACGGCTA |
Para el control negativo se utilizó agua grado molecular y como control positivo las cepas de B. megaterium ATCC®14581 y B. licheniformis ATCC®14580 (Chiluisa-Utreras et al., 2020).
Siembra de semillas
La siembra de las semillas de tomate riñón (Lycopersicum esculentum Mill.) se realizó en bandejas de germinación de 220 hoyos con turba (Promix GTX) humedeciendo el sustrato y dejando las bandejas bajo condiciones de invernadero. La variedad evaluada fue Pietro (Clause).
Preparación del inóculo
Siguiendo lo descrito por Acurio et al. (2020), se elaboró un bioprepa rado sólido para cada cepa. La aplicación se realizó mediante aspersión utilizando una bomba de mochila, la solución aplicada tuvo una concentración bacteriana final de 1x106 UFC/mL.
Se realizó la aplicación de 5 tratamientos, descritos en la Tabla 2.

Grosor del tallo
Para medir el grosor del tallo, se tomaron 20 plántulas al azar de cada bandeja de germinación y mediante el uso de un pie de rey se procedió a tomar las mediciones del diámetro del tallo en milímetros, las mediciones se realizaron semanalmente desde la segunda semana después de la siembra, hasta la semana que las plántulas estuvieron listas para el trasplante (semana 5).
Eficiencia fotosintética
Se tomaron 10 plántulas indistintamente de cada unidad experimental, mediante el equipo OS-30p+Chlorophyll Fluorometer se les realizó la medición de la eficiencia cuántica del fotosistema II (PSII), mediciones que se efectuaron en las semanas 3, 4 y 5.
Materia seca de la parte aérea
De las 20 plántulas seleccionadas en un inicio se pesaron la parte fresca de la planta sin la raíz, el material vegetal se colocó en fundas de papel dentro de una estufa a 70 °C durante 48 horas, a continuación, se colocaron las fundas en un desecador por 24 horas, se tomó los datos de las muestras secas mediante el uso de una balanza analítica.
Resultados y discusión
Identificación molecular de las cepas de Bacillus
En la cepa CT11 los cebadores específicos para el gen phaC, permitieron la amplificación de la secuencia diana, región del genoma que sirve para reconocer especies de B. megaterium
Por otra parte, en la cepa IB10 los cebadores específicos para el gen lchAA, permitió la amplificación de la secuencia de interés, región que permite identificar especies en B. licheniformis.

Figura 1 Electroforesis de PCR de los genes phaC y IchAA. C1: marcador de peso molecular, C2: control negativo gen phaC, C3: control negativo gen IchAA, C4: gen phaC y C6: gen IchAA.
En la cepa IB10 se amplificó el gen IchAA, correspondiente a la secuencia que se utilizó para identificar a la especie Bacillus linchiniformis como se observa en la Figura 1, de acuerdo a lo reportado por Coronel et al. (2016) el gen tiene 2600 pb.
Para la cepa CT11 se amplificó el gen phaC, según reporta Nayak et al. (2013) esta región tiene 900 pb y se utiliza para identificar a la especie Bacillus megaterium.
Tabla 3 Variables evaluadas (Tukey 5 %)
| Tratamientos | Media del diámetro del tallo (mm) ± sd | Eficiencia fotosintética (Fv/Fm) ± sd | Materia seca (g) ± sd | Longitud de raíz (mm) ± sd |
| T1 | 2,63 ± 0,07 a | 2,87 ± 0,23 ab | 0,22 ± 0,00 a | 79,34 ± 1,71 ab |
| T2 | 1,95 ± 0,10 c | 2,89 ± 0,13 ab | 0,16 ± 0,16 b | 80,75 ± 3,88 a |
| T3 | 2,43 ± 0,10 ab | 2,76 ± 0,06 b | 0,15 ± 0,01 b | 80,26 ± 1,91 a |
| T4 | 2,11 ± 0,29 bc | 3,22 ± 0,04 a | 0,15 ± 0,01 b | 81,85 ± 2,62 a |
| T5 | 2,37 ± 0,18 abc | 2,72 ± 0,18 b | 0,11 ± 0,02 c | 73,37 ± 0,47 b |
Nota: sd: desviación estándar; Fv: Fluorescencia variable; Fm: Fluorescencia máxima.
Parámetros de crecimiento
Para incrementar los rendimientos del cultivo de tomate es de vital importancia la obtención de plántulas sanas y con características de vigor (Costales et al., 2007), por esta razón se evaluó longitud de raíz, diámetro de tallo, eficiencia fotosintética y peso seco de plántula (Tabla 3).
Grosor del tallo
En cuanto al grosor del tallo se logró evidenciar que existió diferencia significativa entre los tratamientos con un p valor < 0,01.
En la Tabla 3 se puede observar en base a la prueba Tukey al 5%, que los tratamientos 1, 3 y 5 tuvieron una acción similar sobre la planta, influyendo de manera positiva en cuanto al grosor del tallo.
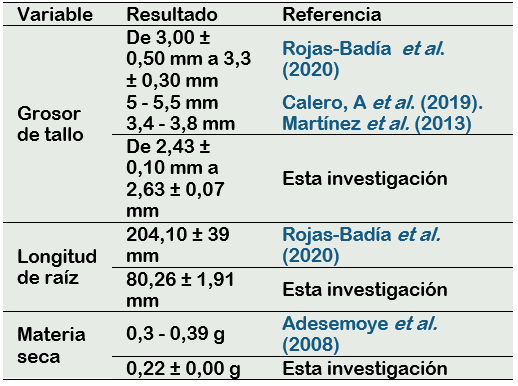
En la investigación realizada por Rojas-Badía et al. (2020), donde se inoculó Bacillus sp. a la siembra de tomate (Solanum lycopersicum), el grosor del tallo de la planta obtuvo un incremento del 90% frente al testigo, reportes que también concuerdan con los presentados por Calero et al. (2019) donde al aplicar RPCV a la siembra y aplicaciones foliares semanales obtuvo un incremento en el diámetro del tallo desde un 30% al 100% en diferentes variedades de Solanum lycopersicum, los resultados obtenidos en esta investigación al aplicar B. megaterium y B. licheniformis a la siembra de tomate el grosor del tallo aumentó hasta en un 10% respecto al testigo.
Según Gómez et al. (2010) uno de los criterios para considerar una óptima calidad en plántulas de tomate a los 25 - 30 días de germinada la semilla es que el grosor del tallo el cual debe estar entre de 2,3 - 5 mm y debido al efecto de las RPCV se evidencia que se alcanza este parámetro de calidad.
Esto puede atribuirse a la capacidad de las bacterias promotoras de crecimiento vegetal, entre ellas especies del género Bacillus, de producir fitohormonas como auxinas y giberelinas que incrementan el crecimiento y grosor de las plántulas, así lo afirma Camelo et al. (2011).
Eficiencia fotosintética
Para la variable eficiencia fotosintética se logró evidenciar que existió diferencia significativa entre los tratamientos con un p valor < 0,05.
Como se puede observar dentro de la Tabla 3 al aplicar la prueba Tukey al 5% se presentan los grupos diferenciados a y b, donde los tratamientos 4, 2 y 1 corresponden al primer grupo ejerciendo un efecto similar en la planta de manera positiva sobre la eficiencia fotosintética.
La fluorescencia del pigmento de clorofila proporciona información sobre la variación que se presentan en la dispersión de calor y la eficiencia fotoquímica, fenómeno que, al ser medido, permite discernir entre los niveles máximos de fluorescencia (Fm) y los niveles mínimos (Fo), valor que representa la supresión del proceso fotoquímico. En plántulas de tomate al inocular B. megaterium semanalmente se presentó un valor mayor de eficiencia fotosintética alcanzando el valor de 3,22 ± 0,04 para la relación Fv/Fm, resultados similares a los obtenidos por Lu et al. (2019), donde al inocular bacterias promotoras de crecimiento a plántulas de Solanum lycopersicum evidenciaron que la eficiencia fotosintética aumenta al doble frente a los controles.
Gracias a la acción de los RPCV, la mayoría de los nutrientes se encuentran a disposición de la planta, lo cual evita que este pase por un proceso de estrés, lo que se evidencia en las elevadas lecturas de Fv/Fm, es decir presentan una mayor eficiencia fotosintética (Gonzales et al., 2008).
Russo y Perkins-Veazie (2010) afirman que cuando se presenta una disminución de la eficiencia fotosintética se puede dar debido a una reducción de nitrógeno en la hoja o en la clorofila y las bacterias promotoras de crecimiento aumentan la disponibilidad de nitrógeno hacia las plantas y a su vez de nutrientes por lo que existe un aumento en la eficiencia fotosintética en las plántulas inoculadas.
Materia seca de la parte aérea
Se presentó diferencia significativa en cuanto al peso de materia seca entre los tratamientos con un p valor < 0,01; donde el tratamiento 1 destaca al aumentar la cantidad de materia seca de la planta (Tabla 3).
Adesemoye et al. (2008) demostraron que al inocular a Solanum lycopersicum L. con B. subtilis a la siembra, la planta de tomate a los 60 días presentó un 36% más de biomasa seca en comparación con el control, Chauhan et al. (2014) reportaron que la cepa CKT1, identificada molecularmente como B. licheniformis, aumentó en un 63,50% el peso seco de Solanum lycopersicum L. En la presente investigación se evidenció que la inoculación de B. licheniformis a la siembra de tomate (T1) provocó un aumento del 100% en la biomasa seca de la planta con respecto al testigo (T5), resultado superior a los reportados en otras investigaciones.
Como lo mencionan Corrales et al. (2017), esto puede deberse a que las rizobacterias promotoras de crecimiento pueden solubilizar nutrimentos que se encuentran inmovilizados y en baja disposición en el suelo o sustratos, como es el caso del fósforo, lo que mejora la asimilación de estos nutrientes hacia la planta, dando como resultado una mayor a cantidad de biomasa.
Longitud de raíz
En cuanto a longitud de raíz existió deferencia significativa entre los tratamientos con un p valor < 0,05; en la Tabla 3 se puede evidenciar en base a la prueba Tukey al 5%, que todos los tratamientos presentan un comportamiento parecido sobre la planta y causan un efecto positivo sobre la longitud de la raíz a excepción del tratamiento 5, el cual corresponde al testigo.
En la investigación realizada por Chauhan et al. (2014), se afirma que en plantas de tomate inoculadas con diversas cepas de Bacillus como B. licheniformis, B. vallismortis, B. subtillis, aumentan la longitud de la raíz desde un 10% al 100% en comparación con las plantas que no han recibido ningún tratamiento, de igual manera Martínez et al. (2013) Evidenció que al adicionar especies del género Bacillus como B. megaterium, B. firmus y B. subtilis sobre la germinación y desarrollo de plántulas de tomate, la longitud de la raíces aumenta desde un 20% al 50% frente a las plántulas que solo se las trató con agua, los resultados presentados en este estudio manifiestan que al inocular B. megaterium y B. licheniformis a las plántulas de tomate semanalmente aumentó en un 11% respecto al testigo.
Según Ribaudo et al. (2006) y Saleem et al. (2007), el ácido indolacético (AIA) absorbido tanto por las semillas como por las raíces de las plántulas, podría incitar la acción de la enzima 1-aminociclopropano-1-carboxilato sintetasa (ACC), la cual es parte de la síntesis del etileno y este en bajas concentraciones incrementa el crecimiento de los pelos radicales de las plantas inoculadas, aumentando así la superficie radicular que favorece a una mayor absorción de nutrientes y esto se traduce a una mayor longitud de raíces.
Comparación con estudios previos
No todas las variedades se adaptan a una región, por tal razón, es necesario que se conozca resultados de estudios de comportamientos agronómicos de las variedades, esto permitirá definir qué tipo de variedades presentan mejores características, resultados de producción y rendimiento para cada zona de un país (INIAP, 2009), esto puede explicar la variabilidad de los resultados en los estudios (Tabla 4).
Por otro lado, la expresión de genes implicados en el crecimiento y desarrollo vegetal varía en función de los cambios ambientales, los cuales modifican dicha expresión génica del organismo (Molina-Romero et al, 2015).
Conclusiones
Los resultados obtenidos en plántulas inoculadas con los microorganismos eficientes (Bacillus licheniformis y Bacillus megaterium) fueron superiores en comparación con los valores del tratamiento testigo. Bacillus licheniformis incrementó significativamente el grosor del tallo y la cantidad de biomasa seca de las plántulas de tomate riñón. En cuanto a la eficiencia fotosintética expresada en Fv/Fm, esta aumentó significativamente debido a la inoculación semanal de Bacillus megaterium. Las aplicaciones de Bacillus licheniformis y Bacillus megaterium semanalmente aumentaron significativamente la longitud de las raíces de las plántulas de tomate.
Se recomienda estudiar la aplicación de los dos microorganismos de manera combinada y en condiciones de invernadero o campo abierto.

