











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
1. Introducción
A pesar de los intentos por retardar la maduración postcosecha de las frutas mediante el almacenamiento a temperaturas bajas, son inevitables los daños que puedan sufrir si se desconocen las condiciones adecuadas de almacenamiento. El almacenamiento en frío sigue siendo el método de conservación más eficaz que se utiliza ampliamente para el almacenamiento postcosecha; el cual podría reducir la tasa metabólica, retrasar la senescencia y mantener la calidad de los vegetales. Sin embargo¸ para algunas frutas y verduras que crecen en los trópicos y subtrópicos, el almacenamiento en frío puede causar daños (Carvajal et al., 2015). Diversos estudios señalan que la calidad sensorial de una fruta almacenada a bajas temperaturas depende de los cambios de ciertos componentes fisicoquímicos y determinados metabolitos. Estos cambios se han observado en diferente frutas como la manzana, donde el almacenamiento a bajas temperaturas afectó su calidad sensorial por una disminución del contenido de ácido málico y de materia seca (Jan & Rab, 2012); en el sapote mamey, temperaturas de almacenamiento entre 10 °C y 15 °C afectaron las características sensoriales como el color y dureza (Díaz-Pérez et al., 2000); en el tomate la biosíntesis del ácido ascórbico fue mayor a 10 °C respecto a 5 °C (Tsaniklidis et al., 2014); en Achras sapota ciertos componentes fisicoquímicos como la acidez, los azúcares reductores y el color de su pulpa fueron afectadas cuando se almacenaron a 4 °C respecto a los almacenado a 16 °C (Vargas et al., 2005), en la palta almacenada a 13 °C se produce un pardeamiento de la pulpa (Woolf et al., 2020), en el melocotón y la nectarina almacenados a 8 °C presentaron un pardeamiento de la pulpa, sin sabor y poca jugosidad (Zhang et al., 2020), caso contrario a la pitahaya roja donde el almacenamiento a 8 °C reportó una disminución en la pérdida de peso, un mayor contenido de acidez titulable, azúcares solubles y de fenoles totales respecto a menores temperaturas (Sheng et al., 2021).
Un adecuado almacenamiento en frutos tropicales como la lúcuma (Pouteria lúcuma), son necesarios a fin de conservar sus caracterizas sensoriales y propiedades funcionales. El Perú es el principal productor y exportador a nivel mundial de esta fruta (SIICEX, 2020), la cual es utilizada como ingrediente en postres, productos de confitería, helados entre otros. La lúcuma es conocida en diferentes países de Sudamérica, historiadores señalan que se consume desde la época pre-inca, por las diversas representaciones encontradas en cerámicos y textiles de la cultura mochica (Villanueva, 2001); con un color de pulpa que varía desde amarillo a amarillo-naranja, de sabor y aroma agradable (Inga et al., 2019). Además, posee propiedades funcionales atribuidos a sus compuestos fenólicos, carotenoides, tocoferoles, fitosteroles, terpenoides, que le confieren capacidad antioxidante (Fuentealba et al., 2016; García-Ríos et al., 2020; Aguilar-Galvez et al., 2021) y a su actividad hipoglucemiante, donde ha mostrado inhibición de las enzimas α-amilasa y α-glucosidasa, implicadas en la hiperglucemia que deviene en la diabetes tipo II (Fuentealba et al., 2016; Aguilar-Galvez et al., 2021). La inexistencia de estudios sobre las condiciones de almacenamiento dificulta un manejo adecuado que eviten cambios o daños en la lúcuma. Por lo tanto, el objetivo de la presente investigación fue evaluar el efecto de las condiciones de almacenamiento en los principales componentes fisicoquímicos, en los metabolitos primarios y secundarios, en la capacidad antioxidante e hipoglucemiante durante la maduración postcosecha de la lúcuma.
2. Materiales y métodos
Fruta. La lúcuma fue recolectada en estado de madurez fisiológica (MF, color amarillo debajo de los sépalos), del biotipo Dos marrón del vivero Topará del Fundo Huayquina, de la provincia de Chincha, departamento de Ica, Perú, las que se dejaron almacenadas hasta alcanzar la madurez organoléptica (MO, ablandamiento de la pulpa), bajo dos condiciones de almacenamiento ambientales (CA, 29 ± 2 °C y 70 ± 2 %HR) y cámara climatizada (CC, 15 ± 1 °C y 90 ± 0,5 %HR). En la Figura 1 se muestra la lúcuma en los dos estados de madurez y en las dos condiciones de almacenamiento.
Metodología experimental. Se analizaron 48 lúcumas (unidades biológicas), para las pruebas no destructivas como pérdida de peso, calibre, color de epicárpio y pulpa, y tasa de respiración, siendo el peso promedio de cada lúcuma de 277 ± 50 g. El muestreo durante el almacenamiento se realizó los días 0, 2, 5, 7, 9 y 12. Para pruebas destructivas como los análisis fisicoquímicos, metabolitos secundarios y propiedades funcionales se analizaron 12 unidades biológicas. Para el análisis de metabolitos primarios por GC-MS; se tomaron 60 unidades biológicas, realizándose el muestreo en los diferentes estados de madurez a través de biopsias (Pedreschi et al., 2014). Para los análisis de metabolitos, capacidad antioxidante e hipoglucemiante las muestras tomadas fueron liofilizadas y almacenadas a -80 °C.
Estándares y Reactivos Químicos. Estándar interno (polares): Kit de ácidos orgánicos 47264, Kit de azúcares CAR10, Kit de azúcares-alcohol 47266 (Supelco, USA), Myo-inositol I5125, Ácido ascórbico 261556 (Sigma, USA), kit de aminoácidos NCI0180 (Thermo Scientific, USA); Acarbosa, Alfa-amilasa (Tipo VI-B> 10 unid/mg), Alfa-glucosidasa, Almidón Soluble, Trolox (Sigma-Aldrich, USA); 1,4 - Dithiothreitol (DTT), Folin-Ciocalteu (Merck®, Alemania).

Figura 1 Pouteria lucuma en estado de madurez fisiológica (A), en estado de madurez organoléptico almacenado en cámara climatizada (B) y en estado de madurez organoléptico almacenado en ambiente (C).
Análisis de algunos componentes fisicoquímicos. La materia seca, acidez titulable, pH, sólidos solubles, fibra dietaría y almidón fueron evaluados de acuerdo con el método descrito por la AOAC (2007). Los azúcares reductores fueron medidos por el método del ácido 3,5-dinitrosalicílico (DNS), recomendado por Miller (1959). La determinación de los minerales calcio (Ca), potasio (K) y magnesio (Mg) se realizó en espectrofotómetro de absorción atómica (Genesys 200, Fisher Scientific, Schaumburg, Illinois, USA) usando estándares de tales elementos (Chapman & Pratt, 1973). La medida de color en la pulpa y epicarpio se realizó por el método CIELAB usando el colorímetro Minolta (Model R-400/410, Japan), tomándose las medidas L, a* b*, C, °h. La pérdida de peso se determinó mediante el pesado de la lúcuma durante el almacenamiento, el porcentaje de pérdida se calculó por la diferencia de pesos respecto al peso inicial de la fruta. La determinación de la tasa de respiración se realizó por el método reportado por Fonseca et al. (2002) con algunas modificaciones con un analizador de gases (MOCON, mod 7500, series 0910ED119); La tasa de respiración se calculó con la siguiente fórmula:

Dónde: V= volumen
La firmeza se determinó mediante el uso de presionómetro Fruit Pressure Tester. (model FT 327 con un rango de 0.5 -13 kg-f), los resultados se expresaron en Newton.
Determinación de metabolitos primarios y secundarios
Azúcares, azúcares alcohol, ácidos orgánicos, aminoácidos. Para la determinación de azúcares, azúcares alcohol, ácidos orgánicos y aminoácidos, se siguió el método descrito por Pedreschi et al. (2014). Para la identificación y cuantificación se utilizó un cromatógrafo de gases Trace 1310 (Thermo Scientific, Rodano, Milan, Italy) acoplado a un espectrómetro de masa triple Quadrupolo modelo TSQ 8000 Evo (Thermo Scientific, USA), empleando una columna HP-5MS UI (0,25 µm, 30m x 0,25 mm ID). Las condiciones de la corrida cromatográfica fueron la temperatura del inyector a 220 °C, la interfase a 280 °C, flujo de Helio a 1 mL/min, fuente a 230 °C, el quadrupolo a 150 °C, volumen de inyección 0,5 µL. Los componentes minoritarios fueron detectados en modo splitless, en una corrida de 50 °C por 1 minuto y luego elevada a razón de 10 °C/min hasta 310 °C y mantenido por 13 minutos. Los componentes mayoritarios fueron detectados en modo Split 1:150, en una corrida a 120 °C por 1 min y luego elevada a razón de 10 °C/min hasta 300 °C y mantenido por 6 minutos. Se realizó curvas de calibración de azúcares, azúcares alcohol, ácidos orgánicos y aminoácidos. Para la cuantificación de los metabolitos polares se utilizó el método del estándar interno, determinándose la concentración relativa, aplicándose la siguiente fórmula:
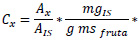
Donde, Cx es la concentración de analito en mg/g ms de la lúcuma, Ax es el área de analito, AIS es el área estándar interno.
Determinación de ácido L-ascórbico. Se aplicó el método descrito por Klimczak & Gliszcznska-Swiglo (2015) con modificaciones. Se pesó 1 g de lúcuma liofilizada y se le adicionó 5 mL de ácido metafosfórico al 10% a 4 °C, sin romper la cadena de frio y en oscuridad se mantuvo en agitación por 10 minutos en agitador magnético, luego se centrifugó por 15 min a 3980 g y 4 °C en centrifuga refrigerada, y se filtró. Luego se realizó la reacción de una fracción del extracto con DL-dithiothreitol (DTT) al 5% para reducir el dehidroascórbico a ácido L-ascórbico. Ambas fracciones, extracto y extracto reducido, fueron filtradas a través de un filtro de 0,22 μm previo al análisis por UPLC. La cuantificación se realizó mediante UPLC-DAD (Waters®, modelo ACQUITY UPLC H-Class, Ireland) con una columna ACQUITY UPLC BEH C18 (1,7 µm, 100 x 2,1 mm DI). La corrida cromatográfica se realizó de acuerdo con lo descrito por Aguilar-Galvez et al. (2021). Los resultados fueron expresados en mg de L-ascórbico/100 g de materia seca.
Determinación de compuestos fenólicos totales. Fueron medidos de acuerdo con el método propuesto por Singleton & Rossi (1965). La extracción de los compuestos fenólicos se realizó de acuerdo con Ma et al. (2004). El contenido de compuestos fenólicos se expresó en miligramos de ácido gálico equivalente por gramo de muestra en materia seca (mg AGE/g ms).
Determinación del contenido total y perfil de Carotenoides. El contenido total de carotenoides se determinó de acuerdo con lo descrito por Aguilar-Galvez et al. (2021), se expresó en mg β-caroteno/g ms; y para la determinación del perfil de carotenoides una fracción fue sometida a saponificación de acuerdo con lo propuesto por Andre et al. (2007) y Aguilar-Galvez et al (2021). Los carotenoides fueron identificados por comparación de los tiempos de retención y espectros de absorción (330-500 nm) con datos reportados en la literatura.
Capacidad antioxidante e hipoglucemiante in vitro
Capacidad antioxidante hidrofilica in vitro. Se aplicó el ensayo TEAC (Trolox equivalent antioxidant capacity) recomendado por Arnao et al. (2001) usando el reactivo ABTS [(2,2’-azino-bis(3-etilbenzotiazolina-6-ácido sulfó-nic)]. Los extractos se obtuvieron con lo propuesto por Wu et al. (2004) con modificaciones. Los resultados fueron expresados en μmol de trolox equivalente (TE)/g muestra ms.
Capacidad hipoglucemiante in vitro. Se pesó 0,5 g de pulpa de lúcuma liofilizada, se le adicionó 25 mL de acetona al 80%, luego se agitó por 90 minutos en agitador magnético, luego se centrifugó a 2397 g por 10 minutos. El sobrenadante recuperado se concentró a vacío hasta 10 mL, a 40 °C en rotavapor. Al extracto obtenido se le ajustó el pH a 7,0±0,05 en potenciométro; y luego se distribuyó 1 mL del extracto en eppendorf y se almacenó a -20 °C hasta el momento del análisis. Para la medida de la capacidad hipoglucemiante se aplicó el método descrito por Gonzalez-Muñoz et al. (2013) y en la determinación del IC50 (concentración inhibitoria del 50% de una sustancia) se ensayó con 6 diferentes diluciones del extracto de lúcuma.
Tabla 1 Cambios en los componentes de la lúcuma almacenada en condiciones ambientales (CA) y en cámara climatizada (CC)
| Contenido | Madurez Fisiológica | Madurez Organoléptica CA | Madurez Organoléptica CC |
| Materia Seca | 35,51±2,50ab | 37,48±2,27aA | 33,23±2,33bB |
| % Acidez titulable (g ac. Cítrico/100 g ms) | 0,76±0,13b | 0,47±0,03aA | 0,43±0,03aA |
| pH | 5,99±0,14b | 5,51±0,04aA | 5,72±0,21aA |
| Sólidos solubles (°Brix) | 15,98±1,64a | 22,17±2,76bA | 18,23±1,26bA |
| Almidón (g/100 g ms) | 12,90±3,42b | 6,11±0,71aA | 9,0±1,40aA |
| Azúcares reductores (g glucosa/100 g ms) | 15,19±2,84a | 27,80±5,39bA | 21,74±1,51bA |
| Fibra dietaría (g/100 g ms) | |||
| Fibra Insoluble | 19,15± 2,04a | 22,61±3,85bA | 26,86±3,09bA |
| Fibra soluble | 5,32± 0,76b | 4,32±0,45aA | 3,75±1,25aA |
| Fibra Total | 24,47± 2,33a | 26,93±4,04bA | 30,60±2,70bA |
| Minerales (%) | |||
| Potasio | 0,94±0,07a | 0,99±0,24 aA | 0,91±0,01 aA |
| Calcio | 0,10±0,02 a | 0,08±0,01 aA | 0,08±0,01 aA |
| Magnesio | 0,03±0,00 a | 0,03±0,00 aA | 0,03±0,00 aA |
| Firmeza (N) | 127,49±0,00b | 5,93±1,69aA | 8,66±1,72aA |
Letras minúsculas diferentes en una misma fila indican diferencia significativa (p < 0,05) por efecto del estado de madurez, letras mayúsculas en una misma fila indican diferencias significativas (p < 0,05) por efecto de las condiciones de almacenamiento. Repeticiones n =4 (unidades biológicas).
Diseño experimental y análisis estadístico. Se aplicó una factorial 2K, donde k es 2; siendo un factor la condición de almacenamiento con los niveles: CA y CC; y el otro factor fue estado de madurez con niveles: Madurez fisiológica y Madurez organoléptica (MO). Para el caso de las pruebas físicas como la medida de color, pérdida de humedad, tasa de respiración, se aplicó un diseño factorial con dos factores; siendo un factor la condición de almacenamiento con los niveles: CA y CC; y el otro factor fue tiempo de almacenamiento con 6 niveles: 0, 2, 5, 7, 9 y 12 días. Cada muestra biológica es una repetición.
Análisis estadístico. Se aplicó análisis de varianza (ANVA), si hay significancia se emplea un análisis de múltiples rangos mediante la prueba de Tukey. El análisis estadístico se realizó mediante el programa Statgraphics Centurion 18-X64 USA.
3. Resultados y discusión
Componentes fisicoquímicos relacionados con la maduración postcosecha
El contenido de materia seca fue afectado significativamente (p < 0,05) por la condición de almacenamiento (Tabla 1), estos resultados son similares a lo reportado en lúcuma de diversos biotipos y de diferente estado de madurez fisiológica y organoléptica (Fuentealba et al.,2016; García-Ríos et al., 2020; Aguilar-Galvez et al., 2021). El contenido de acidez (Tabla 1), disminuyó significativamente (p < 0,05) desde 0,76 hasta 0,47 y 0,43 g ácido cítrico/100 g ms en la almacenada a CA y CC, respectivamente, influenciada por el estado de madurez. Sin embargo, García-Ríos et al. (2020) no registró variación significativa en las variedades Seda y Beltrán, alcanzando valores de 0,28 y 0,36 g ac. cítrico/100 g ms. Diferentes investigaciones indican que una diminución de la acidez coincide con el avance de la maduración, por el incremento de la tasa respiración, donde los ácidos orgánicos son el sustrato para la respiración (Ackermann et al., 1992; Wang et al., 1993). El pH presentó una disminución significativa desde 5,99 hasta 5,51 y 5,72 en lúcuma almacenada en CA y CC, respectivamente, estando afectadas solo por el estado de madurez. Esta disminución de la acidez estaría asociada con la desmetilación de los homogalacturanos que durante la maduración crea un incremento de regiones cargadas negativamente, dejando cargado al grupo carboxilo, e influyendo en el pH (Grignon & Sentenac, 1991).
Los sólidos solubles se incrementaron (Tabla 1); siendo afectado significativamente solo por el estado de madurez, otras variedades de lúcuma presentaron valores similares en lúcuma en estado de madurez organoléptica 21,7 y 23,4 °Brix en Beltrán y Seda (García-Ríos et al.,2020); 29,49 y 32,1 °Brix en variedad Trompito y María Belén (Alegre & Ticse, 2017). El contenido de almidón disminuyó significativamente (p < 0,05) desde 12,9 hasta 6,1 y 9,0 g/100 g ms en CA y CC, respectivamente (Tabla 1), afectado por el estado de madurez; otras variedades como Beltrán y Seda reportaron valores superiores de 15,6 y 11,7 g/100 g ms en MO (García-Ríos et al.,2020); este contenido de almidón alcanza un valor máximo en la etapa de madurez fisiológica y luego es casi completamente degradado con un aumento simultáneo de azúcares cuando alcanza la madurez organoléptica (Mesa et al., 2016). El contenido de azúcares reductores se incrementó significativamente (Tabla 1), por efecto solo del estado de madurez. Por su parte García-Ríos et al. (2020) reportó similares contenidos de azúcares reductores en MO de 27,2 y 23,2 g glucosa/100 g ms en variedad Beltrán y Seda, respectivamente. La fibra dietaría total (FDT) varió significativamente (p < 0,05) desde 24,5 hasta 26,9 y 30,6 g/100 g ms en CA y CC, respectivamente, siendo significativo en la lúcuma almacenada en CC. La mayor proporción por la FDT corresponde a fibra insoluble (FDI) mostrando un mayor incremento durante el almacenamiento en CC. Otros estudios en lúcuma en madurez organoléptica, registraron valores de FDT menores a los encontrados en la presente investigación, 24,2% en la variedad Seda y Beltrán (García-Ríos et al., 2020); 15, 15,18 y 13,7 en las variedades Beltrán, Trompito y María Belén, respectivamente (Alegre & Ticse, 2017). El contenido de FDT que aporta la lúcuma es apreciable, comparado con otros frutos, como la sapodilla (17,3), sapote mamey (17,2); mango maduro (8,2), mango verde (10,6), papaya verde (23,7) y papaya madura (14,0) todos en porcentaje respecto a la materia seca (Mahattanatawee et al., 2006). La firmeza de la lúcuma es afectada por el estado de madurez (p < 0,05), decreciendo significativamente desde 127,5 N hasta 5,9 y 8,7 N en lúcumas almacenas en CA y CC, respectivamente. El ablandamiento del fruto es un proceso de modificaciones en la estructura de la pared celular donde se presenta despolimerización de glicanos y solubilización de pectina (Fruk et al., 2014); así como a la degradación del almidón en sólidos solubles, los cuales influyen sobre la pared primaria (Jiang et al., 2019). Respecto al color de pulpa (Figura 2), no se afectó por la condición de almacenamiento, teniendo una tonalidad amarilla-naranja por la presencia de carotenoides del grupo de las xantofilas como la neoxantina, violaxantina, zeaxantina e isómeros de luteína (Fuentealba et al., 2016; Aguilar-Galvez et al., 2021).
La medida de color del epicarpio L* (Figura 3A), °h (Figura 3B), C (Figura 3C), a* (Figura 3D), b* (Figura 3E), muestran cambios significativos (p < 0,05) en ambas condiciones de almacenamiento, siendo similares estos valores luego de los 12 días de almacenamiento, a excepción de los parámetros a* y °h donde se observó diferencias en las dos condiciones de almacenamiento, el cálculo de ΔE de la lúcuma fueron de 5,43 ± 0,51 y 9,37 ± 0,87 en CA y CC respectivamente, mostrando una diferencia significativa entre ambas condiciones de almacenamiento. Los resultados indicaron un cambio de la tonalidad del epicarpio desde verde a verde amarillo; siendo más notorio en la lúcuma almacenada en CC; similares a la reportado en lúcumas de variedad seda (García-Ríos et al., 2020). Con relación al peso (Figura 4), se observó una mayor pérdida en la lúcuma almacenada en CA (10,8%) respecto a la almacenada en CC (3%), mostrando que existe un efecto significativo (p < 0,05) de la condición y tiempo de almacenamiento, así como el efecto conjunto de ambos factores.
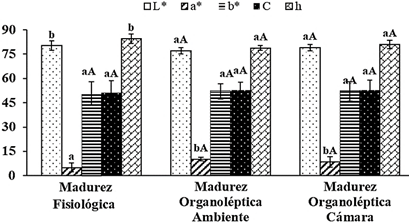
Figura 2 Promedio de L*, a*, b*, C y h de la pulpa fresca de lúcuma almacenada: condiciones ambientales (CA) y en cámara climatizada (CC). Letras mayúsculas diferentes para un mismo color de barra indican diferencias significativas (p < 0,05) por efecto del tiempo de almacenamiento. Letras minúsculas diferentes para un mismo color de barra indican diferencias significativas (p < 0,05) por efecto del estado de madurez.
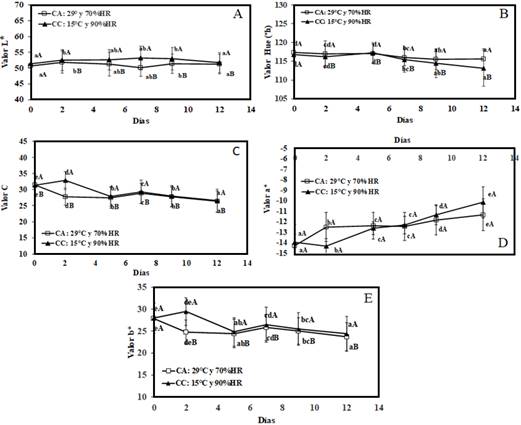
Figura 3 Medida de color del epicarpio, valor L* (A), valor Hue (h) (B), valor C (C), valor b* (D), valor a* (E) de lúcuma almacenada: condiciones ambientales (CA), y en cámara climatizada (CC). Letras mayúsculas diferentes indican diferencias significativas (p < 0,05) por efecto del tiempo de almacenamiento. Letras minúsculas diferentes en sentido vertical indican diferencias significativas (p < 0,05) por efecto del tipo de almacenamiento.
Tabla 2 Contenido de metabolitos de lúcuma almacenada en condiciones ambientales (CA) y en cámara climatizada (CC)
| CONTENIDO | MF | CA | CC |
| Azúcares (mg/g ms)1 | |||
| Fructosa | 25,78 ± 11,09a | 161,43 ± 81,36bA | 206,25 ± 28,13bbA |
| Glucosa | 80,59 ± 22,46a | 152,12 ± 44,48bA | 236,21 ± 39,99bB |
| Sacarosa | 121,58 ± 28,88a | 130,83 ± 36,56aB | 113,90 ± 52,67aB |
| Azúcares alcohol (mg/g ms)1 | |||
| Myo-Inositol | 5,75 ± 1,61a | 7,97 ± 2,27bA | 8,35 ± 1,75bB |
| Manitol | 0,06 ± 0,03a | 5,4 ± 3,68bA | 4,17 ± 1,97bB |
| Ácidos Orgánicos (mg/g ms)1 | |||
| Ac. Cítrico | 0,01 ± 0,01a | 0,02 ± 0,01a | ND |
| Ac. Quínico | 0,05 ± 0,04a | 0,03 ± 0,02bA | 0,03 ± 0,01bA |
| Ac. Málico | 0,40 ± 0,13a | 0,25 ± 0,19bA | 0,16 ± 0,09dA |
| Ac. Ascórbico2 | 0,48 ± 0,04a | 0,27 ± 0,13bA | 0,19 ± 0,07bA |
| Aminoácidos (ug/g ms)1 | |||
| Alanina | 17,5 ± 9,80a | 160,48 ± 126,20bA | 35,78 ± 10,82cB |
| Valina | 1,01 ± 1,04a | 21,24 ± 16,30bA | 4,46 ± 4,02bB |
| Tirosina | 33,7 ± 8,72a | 28,35 ± 25,06aA | 23,34 ± 24,24aA |
| Ac. Glutámico | 100,18 ± 95,97a | 4,53 ± 4,49bA | 6,07 ± 3,69bA |
| Ac. Aspártico | 12,68 ± 11,20a | 0,99 ± 0,55bA | 0,41 ± 0,47bA |
| Treonina | 4,85 ± 2,84a | 9,4 ± 6,07bA | 4,91 ± 4,39bA |
| Comp Fenólicos Tot. (mg AGE/g ms)2 | 55,33 ± 2,36a | 1,83 ± 1,83bA | 0,98 ± 0,11bA |
| Carotenoides Tot. (mgβ caroteno equiv./g ms)2 | 0,22 ± 0,03a | 0,17 ± 0,02bA | 0,14 ± 0,02bA |
| µgβ caroteno/g ms | 3,09 ± 1,31a | 2,73 ± 0,95 bA | 1,88 ± 0,52 bA |
1. Metabolitos determinados por GC-MS (n = 30) expresados en concentraciones relativas, 2. Metabolito determinado por espectrofotometría (n = 4) expresados en mg analito/g ms, MF: Madurez fisiológica. Letras minúsculas diferentes en una misma fila indican diferencia significativa (p < 0,05) por efecto del estado de madurez. Letras mayúsculas en una misma fila indican diferencias significativas (p < 0,05) por efecto de las condiciones de almacenamiento. ND: No determinado. La variabilidad es por las unidades biológicas.
Esta pérdida de peso es proporcional a la gradiente de humedad relativa entre el ambiente y la fruta (Wills & Golding, 2016); existiendo una mayor gradiente en las lúcumas almacenadas en CA respecto a la almacenada en CC. La tasa de respiración (Figura 5) fue afectada por las condiciones de almacenamiento, tiempo de almacena miento y por el efecto conjunto de ambos factores (p < 0,05), llegando alcanzar valores máximos de 86,8 y 91 mL CO2/ kg h en el día 9 en CA y CC, respectivamente, describiendo la lúcuma un comportamiento de una fruta climatérica (Watada et al., 1984). Los valores registrados en el pico climatérico lo consideran como un fruto de una tasa de respiración extremadamente alta, similar al espa rrago, brócoli, champiñones entre otros (Saltveit, 2016).
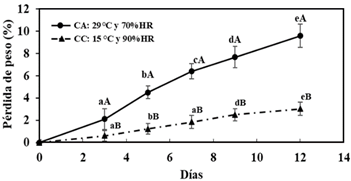
Figura 4 Pérdida de peso de lúcuma almacenada: condiciones ambientales (CA) y en cámara climatizada (CC). Letras minúsculas en una misma línea indican diferencias significativas (p < 0,05) respecto al tiempo, letras mayúsculas diferencias significativas (p < 0,05) respecto a la condición de almacenamiento.
Evolución de metabolitos primarios y secundarios
Respecto al contenido de azúcares de la lúcuma (Tabla 2), se observó un incremento significativo (p < 0,05) de la fructosa y de la glucosa, mientras que la sacarosa presentó variaciones no significativas; similar tendencia lo mostró la lúcuma de variedad Leiva 1, donde el contenido de fructosa se incrementó desde 33,9 hasta 98,7 mg/g ms, la glucosa varió desde 59,4 hasta 170,9 mg/g ms y la sacarosa presento un incremento no significativo desde 32,2 hasta 36,2 mg/g ms (Fuentealba et al., 2016).
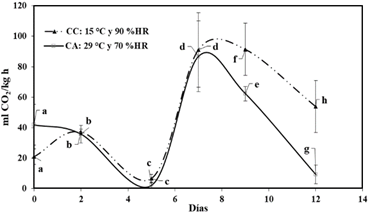
Figura 5 Tasa de respiración de lúcuma almacenada: condiciones ambientales (CA) y en cámara climatizada (CC).
Los azúcares alcohol como myo-inositol y manitol se incrementaron significativamente (Tabla 2); tendencia similar a lo registrado en lúcuma de variedad Leiva 1 (Fuentealba et al., 2016); el myo-inositol es importante por su participación como precursor en la síntesis del ácido ascórbico, a partir de la glucosa (Loewus & Murthy, 2000). Los ácidos orgánicos hallados fueron el ácido cítrico, quínico y málico, otros estudios en lúcuma identificaron adicionalmente el succínico y tartárico (Fuentealba et al., 2016; García-Ríos et al., 2020; Aguilar-Galvez et al., 2021).
Respeto al contenido de ácido ascórbico sólo fue afectado por el estado de madurez (Tabla 2), observándose una disminución significativa (p < 0,05) desde 0,48 hasta 0,27 y 0,19 mg/g ms en CA y CC respectivamente, resultado similares a lo registrado en lúcumas variedad Leiva 1 donde disminuyó desde 0,47 hasta 0,19 mg/g ms (Fuentealba et al., 2016), valores superiores en lúcuma de MO fueron reportados por García-Ríos et al. (2020) de 0,68 y 0,58 mg/g ms en lúcumas variedades Beltrán y Seda. Esta dis minución puede atribuirse a que el ácido ascórbico podría haberse utilizado como cofactor de enzimas, o como an tioxidante contra especies reactivas de oxígeno, así como, también pueden actuar como cofactor en la síntesis de etileno (Gallie, 2013). Respecto a los aminoácidos (Tabla 2); los ácidos Glu, Asp y Thr fueron afectadas sólo por el estado de madurez; la variación del contenido de ami noácidos muestra diversas tendencias, relacionadas con el anabolismo y catabolismo que se activan durante la ma duración postcosecha de la lúcuma, así como, participan en la síntesis de enzimas que intervienen en la maduración (Zhang et al.,2010). El contenido de compuestos fenólicos totales, fueron afectados sólo por el estado de madurez (Tabla 2), observándose una disminución significativa desde 55,33 hasta 0,98 y 1,83 mg AGE/ g ms en la lúcuma almacenada en CC y CA, respectivamente; similar tenden cia fue reportado en lúcuma variedad Leiva 1 variando desde 45,3 hasta 0,7 mg AGE/g ms (Fuentealba et al., 2016); otros estudios reportan contenidos superiores en lúcuma en estado de madurez organoléptica 2,4 mg AGE/g ms (García-Ríos et al., 2020) y 11,4 mg AGE/g ms (Pinto et al., 2009). El contenido de carotenoides totales sólo fue afectado por el estado de madurez, observán dose una disminución significativa (p < 0,05) desde 0,22 hasta 0,17 y 0,14 mg β caroteno/g ms en CA y CC, res pectivamente (Tabla 2). Otros estudios en lúcuma repor tan valores de carotenoides cercanos como la variedad Leiva 1 desde 0,25 hasta 0,23 (Fuentealba et al., 2016), la variedad Dos marron de 0,148 en madurez fisiológica (Aguilar-Galvez et al., 2021) y García-Ríos et al. (2020) reportaron 0,25 y 0,30 para las variedades Seda y Beltrán respectivamente, todos expresados en mg β caroteno/g ms. La variabilidad de los resultados mostrados en las Tablas 1 y 2, se atribuye a la heterogeneidad de las unidades biológicas (una lúcuma es una unidad experimental), durante el proceso de maduración; similares variabilidades fueron reportados por Fuentealba et al (2016) en lúcuma de diferentes estados de madurez.
El estudio del perfil de carotenoides (Figura 6) se aprecia que no cambian durante la maduración postcosecha en las dos condiciones de almacenamiento más si en las áreas, lo cual indicaría que no hay un cambio importante en la síntesis de carotenoides desde la madurez fisiológica hasta la madurez organoléptica; los 10 primeros picos per tenecen a la familia de las xantofilas y sólo dos picos, a los carotenos. Por comparación de los espectros de absor ción, el pico 1 corresponde a auroxantina (Inbaraj et al., 2006), los picos 2 y 3 corresponden a trans-neoxantina y trans-violaxantina, respectivamente (Simonovska et al., 2013). El pico 4 fue el mayoritario en todos los casos y de acuerdo a su perfil de absorción se puede atribuir a un isómero de la luteína (Kao et al., 2012), el pico 5 se identi ficó como neoxantina por su espectro de absorción y tiempo de retención con el estándar de este carotenoide, los picos 6 y 7 no fueron identificados, los picos 7 y 8 se identificaron como trans zeaxantina y 9-o-9’ cis luteína (Kao et al., 2012), el pico 10 corresponde al 9 o 9´-cis-β-criptoxantina, el pico 11 correspondería al 9 o 9´-cis-β-caroteno y el pico 12 el trans-β-caroteno (Rojas-Garbanzo et al., 2011). Los perfiles obtenidos en la presente investi gación concuerdan con lo reportado por Aguilar-Galvez et al. (2021); mientras que Fuentealba et al. (2016) y García-Ríos et al. (2020), no reportaron a la auroxantina y al caroteno 9 o 9´-cis-β-caroteno. El contenido de β caroteno, presentó una variación no significativa.
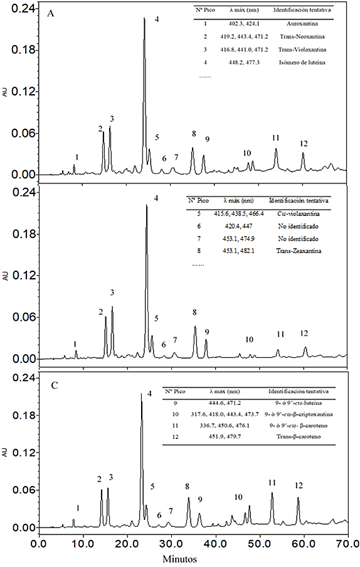
Capacidad antioxidante hidrofílica in vitro
La capacidad antioxidante hidrofílica indica que sólo fueron afectados por el estado de madurez, disminuyendo desde 1415 ± 490,26 hasta 9,60 ± 4,96 y 10,12 ± 6,06 μmol TE/g ms en CA y CC, respectivamente; Fuentealba et al. (2016) registró la misma tendencia variando desde 1096,9 hasta 4,8 μmol TE/g ms en variedad Leiva 1; Aguilar-Galvez et al. (2021) reportó 992,7 μmol TE/g ms en lúcuma en MF; García-Ríos et al. (2020) registró 19,3 y 17,3 μmol TE/g ms en MO de las variedades Beltrán y Seda. Esta disminución de la capacidad antioxidante muestra una correlación altamente significativa (p < 0,01), con el contenido de compuestos fenólicos totales (r = 0,9295), así como con el contenido de ácido ascórbico (r = 0,7711). Por su parte Arnao et al. (1996) señala que tanto los compuestos fenólicos, así como el ácido ascórbico contribuyen en la capacidad antioxidante.
Actividad hipoglucemiante in vitro
La actividad hipoglucemiante muestra que sólo es afectada significativamente por el estado de madurez. La IC50 de la α-glucosidasa (Tabla 3) se incrementó significativamente (p < 0,05), siendo este incremento en más de 250 veces en estado de madurez organoléptico con respecto al de madurez fisiológica. Tendencia similar fue reportado por Fuentealba et al. (2016) en madurez fisiológica 10 µg para lograr el 50% inhibición, mientras que en madurez organoléptica 384 µg ms aprox. para lograr el mismo efecto sobre la α-glucosidasa.
Tabla 3 Valores de concentración inhibitoria media máxima (IC50) de α-glucosidasa de lúcuma almacenada en condiciones ambientales y en cámara climatizada
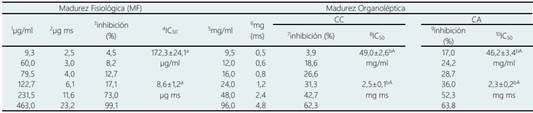
1, 5: Concentración de extracto de lúcuma; 2, 6: Peso de lúcuma; 3, 7, 9: Porcentaje de inhibición de la α-glucosidasa; 4, 8, 10: IC50 expresados en concentración de extracto y peso. Letras minúsculas diferentes en una misma fila indican diferencias significativas (p < 0,05) por efecto del estado de madurez. Letras mayúsculas en una misma fila indican diferencias significativas (p < 0,05) por efecto de las condiciones de almacenamiento. Repeticiones n = 4 (unidades biológicas).
Tabla 4 Valores de concentración inhibitoria media máxima (IC50), de α-amilasa de lúcuma almacenada en condiciones ambientales y en cámara climatizada
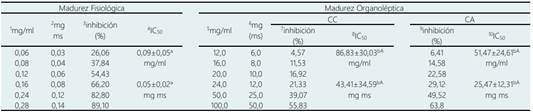
1, 5: Concentración de extracto de lúcuma; 2, 6: Peso de lúcuma; 3, 7, 9: Porcentaje de inhibición de la α-amilasa; 4, 8, 10: IC50 expresados en concentración de extracto y peso. Letras minúsculas diferentes en una misma fila indican diferencia significativa (p < 0,05) por efecto del estado de madurez. Letras mayúsculas en una misma fila indican diferencias significativas (p < 0,05) por efecto de las condiciones de almacenamiento. Repeticiones n = 4 (unidades biológicas).
Pinto et al. (2009) evaluó la lúcuma en estado de MO, si bien no evaluó IC50, se puede inferir de los valores reportados para una dosis aproximada de 31,25 mg/ml registra 50 % de inhibición. Con relación a la inhibición de la α-amilasa (Tabla 4), presentó un comportamiento semejante a la α-glucosidasa, tendencia similar fue reportado por Fuentealba et al. (2016), en donde la lúcuma en madurez fisiológica requiere 0,31 mg ms para lograr el 50% inhibición. Por otro lado, en la presente investigación en lúcuma de madurez organoléptica, necesita una mayor dosis para lograr un efecto inhibitorio del 50% en relación con lo hallado por Fuentealba et al. (2016) de 6,25 mg ms, pero similar a lo reportado por Pinto et al. (2009) de 83 mg/ml. La α-amilasa y la α-glucosidasa están asociadas con la digestión de los carbohidratos y con el metabolismo de la glucosa, la inhibición de estas enzimas ayuda a reducir los niveles de glucosa en la sangre (Pinto et al., 2009).
4. Conclusiones
La condición de almacenamiento en cámara climatizada tuvo un efecto positivo sobre la pérdida de peso durante la maduración postcosecha de la lúcuma, confiriéndole una mejor apariencia físico-sensorial; más no así sobre los cambios de la mayoría de los metabolitos primarios y secundarios, capacidad antioxidante e hipoglucemiante que decrecieron significativamente cuando alcanzaron la madurez organoléptica; a pesar de la disminución de componentes con propiedades funcionales, los valores alcanzados cuando la lúcuma está madura organolépticamente resulta ser ventajosa frente a otras frutas, y su consumo podría ser beneficioso. Por ello, sería importante evaluar diferentes condiciones de almacenamiento de la lúcuma que retarden el proceso de maduración y eviten la perdida de la mayoría de los componentes que le confieren propiedades funcionales.

