











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
1. Introducción
Los granos coloreados del maíz poseen propiedades saludables, principalmente debido a su contenido de antocianinas y carotenoides (Colombo et al., 2021) y las razas nativas de maíz proporcionan cualidades únicas a la cocina peruana (Salvador-Reyes et al., 2021). El consumo de antocianinas es beneficioso para la salud (Chuntakaruk et al., 2021; Cui et al., 2021; Tang et al., 2017; He & Giusti, 2010; Li et al., 2020), una manera de promover la ingesta de este pigmento es a través del desarrollo de variedades de maíz morado reventón; sin embargo, la mejora genética para reunir en una sola variedad caracteres de valor, relacionados a la calidad de grano y productividad, está en su fase inicial.
Los caracteres cuantitativos, se dicen que son poligénicos porque están bajo el control de muchos genes, principalmente de efectos aditivos, responsables de la variación continua (Nilsson-Ehle, 1909; Klug et al., 2017; Zhang et al., 2018), así mismo, tienen una fuerte influencia ambiental, lo que dificulta el estudio de su herencia genética y su mejora.
En el maíz reventón, el volumen de expansión (VE) es el principal carácter (de Oliveira et al., 2020), pero el modo de herencia y el número de genes que controlan su expresión no están del todo dilucidados. Lu et al. (2003) mencionan que el VE está controlado por pocos genes, entre 3 a 5; Dofing et al., (1991), señalan que los efectos genéticos dominantes son los más importantes; Da Silva et al. (2010) y Crumbaker et al. (1949) encontraron un predominio de dominancia parcial y aclaran que el volumen de expansión bajo es parcialmente dominante sobre el volumen de expansión alto, que es recesivo; sin embargo, para la mayoría de autores, genes con efectos genéticos aditivos son los responsables de su herencia y añaden que los efectos genéticos aditivos como dominantes son los más importantes (Pereira & Amaral Júnior, 2001; Arnhold et al., 2010; Coan et al., (2019). Ziegler (2001) menciona que son 4 o 5 genes de efectos mayores y muchos genes de efectos menores los que controlan la herencia de esta característica, lo que significa que el modo de herencia es del tipo cuantitativo. En maíz reventón, el rendimiento de grano (RG), que es un atributo cuantitativo controlado por muchos genes, no es una característica principal, desde que, si un genotipo tiene una mala expansión o mal sabor, no será importante así tenga alto rendimiento (Ziegler, 2001); sin embargo, los genes responsables del RG actúan de manera sobredominante (Da Silva et al., 2010; Pereira y Amaral Júnior, 2001). Ramírez et al. (2020) evaluaron 42 accesiones de libre polinización, 8 variedades élites y un testigo comercial, para condiciones del municipio de Texcoco, Estado de México, el rendimiento promedio fue de 3.38 t/ha, el volumen de expansión (VE) de 2,73 cm³/g y 32,4% porcentaje de granos reventados (PGR); el testigo comercial obtuvo 19,33 cm³/g para VE y 72,5% para PGR (Ramírez et al., 2020). Para condiciones de Turquía, el VE de 12 genotipos comerciales de maíz reventón varió entre 18,50 cm3/g y 35,25 cm3/g (Soylu & Tekkanat, 2007).
En el maíz, los colores del grano más comunes son amarillo, rojo, morado y blanco. Dos son las vías metabólicas responsables de estos colores, la de los carotenoides del color amarillo y la de la antocianina que producen una amplia gama de colores que van desde el naranja-rojo hasta el violeta-azul, este enorme rango de variación depende de modificaciones en su estructura, de co-pigmentos, de iones metálicos, del pH de las vacuolas, la localización de las antocianinas y las formas de las células de la superficie, que contribuyen al color final de órgano o tejido; el blanco se produce por falta de pigmentos en ambas vías (Yoshida et al., 2009; Tanaka et al., 2008). Las antocianinas -cuyo contenido es superior en maíces de granos morados (Suriano et al., 2021)- son solubles en agua, se sintetizan en el citosol y se acumulan en las vacuolas. La biosíntesis de las antocianinas está influenciado por señales intrínsecas ambientales, como la temperatura (Chalker‐Scott, 1999; Aguilar-Hernández et al., 2019) y depende de la acción de tres grupos de genes: i) los genes estructurales, que codifican enzimas como la chalcona sintasa, la chalcona isomerasa, la flavanona 3-hidroxilasa, el flavonoide 3'-hidroxilasa, la dihidroflavonol 4-reductasa, leucoantocianidina oxigenasa, antocianidina reductasa y UDP-glucosa: flavonoide 3-O-glucosil-transferasa (Oh et al., 2021; Holton & Cornish, 1995; Selinger & Chandler, 1999; Pelletier et al., 1997; Harborne & Williams, 2000; Winkel-Shirley, 2002), la malonil-CoA que es un intermedio importante en la síntesis de antocianinas (Peer et al., 2001) y la antocianidina sintasa que cataliza la síntesis de las correspondientes antocianidinas coloreadas (Tanaka et al., 2009;Tanaka et al., 2008), ii) los genes reguladores, que controlan los factores de transcripción, llamados activadores de la transcripción que codifican proteínas responsables de la transcripción de genes estructurales y de los que son responsables de la expresión diferencial de fitohormonas (Meng et al., 2021; Ming et al., 2021) y iii) los genes que se sobre expresan, como el gen Lc que controla el color de la hoja (Xiang et al., 2021). Los genes Pr1, C1 y R1 controlan la síntesis de antocianinas; sin embargo, se requieren otros genes que complementan esta vía metabólica que tiene múltiples pasos. El gen estructural Pr1 codifica la proteína flavonoide 3-hidroxilasa, enzima responsable del color púrpura de la aleurona y el pericarpio (Žilić et al., 2012; Coe et al., 1988); el gen estructural Y1 codifica la fitoeno sintasa, responsable del color amarillo que se acumula en el endospermo (Wurtzel et al., 2012; Coe et al., 1988). Los genes C1 y R1, codifican activadores de la transcripción que regulan varios genes estructurales dentro de la vía de las antocianinas. Además de los genes Pr1, C1 y R1, más de 20 genes, que comprenden genes estructurales y reguladores, deben estar presentes en al menos una dosis del alelo dominante para la síntesis de estos pigmentos, cuando el alelo recesivo r1 o c1 está presente como homocigoto recesivo, no se produce antocianina y el grano es amarillo o blanco, según los genes presentes para la vía de los carotenoides (Coe et al., 1988). Cone (2007), identificó siete genes estructurales c2, chi, pr1, fht1 (F3H), a1, a2, bz1, seis genes reguladores c1, pl1, r1, b1, in1, pac1 y dos genes transportadores de pigmentos hacia las vacuolas, bz2, ZmMrp4. Las pelargonidinas predominan en los granos rojo-rosa y las antocianinas abundan en los granos de color púrpura oscuro (Peniche & Tiessen, 2020) y se detectan a partir de los 21 días después de la floración femenina (DFF), con la máxima acumulación a los 29 DFF (Kim et al., 2020).
El modo de acción de estos genes es variable, por ejemplo, en los resultados de las siguientes autofecundaciones se observan proporciones distintas R1r1/Pr1Pr1/Y1Y1/C1C1 3/4 morados: 1/4 blanco (dominancia completa); R1r1/pr1pr1/y1y1/C1c1 9/16 rojos: 7/16 blancos (epistasis doble recesiva); R1r1/Pr1Pr1/Y1y1/C1C1 12/16 morados: 3/16 amarillo: 1/16 blanco (epistasis dominante simple), lo que evidencia la complejidad en el estudio y la mejora de esta característica (Ford, 2000). La variación en la intensidad de color en los granos morados parece ajustarse más a la expresión de un carácter cuantitativo (QTL) controlado por muchos genes, las herramientas moleculares modernas parecen confirmarlo (Chatham & Juvik, 2021; Sa et al., 2020). Estas evidencias científicas indican que los caracteres, volumen de expansión y antocianina en los granos, tienen modo de herencia compleja y los genes responsables de estos atributos están separados en variedades nativas peruanas, sin embargo, no se conoce cuál será el comportamiento cuando los genes que controlan estos dos caracteres estén en una misma variedad, por ello los objetivos de esta investigación son (1) analizar el modo de segregación de volumen de expansión (VE), expansión de granos (EG), pericarpio negro (PN) y rendimiento de grano (RDG) en 256 líneas S1 y (2) seleccionar líneas S1 de maíz reventón morado sobresalientes para VE y PN.
2. Materiales y métodos
2.1 Ubicación. Los datos para el análisis provienen de tres experimentos, uno de Huánuco y dos de La Molina. En Huánuco fue instalado en el campo de la Universidad Nacional Hermilio Valdizán, Latitud: 09°55′15″, Longitud: 76°18′35″ y altitud de 1986 msnm, los otros dos experimentos fueron instalados en La Molina, en el campo de la Universidad Nacional Agraria La Molina, Latitud: 12°04′55″, Longitud: 76°56′53″ y altitud de 241 msnm.
2.2 Material biológico. El material genético estuvo conformado por 256 líneas S1, pertenecientes a la primera generación de autofecundación, cuyo origen es el siguiente: dos poblaciones nativas peruanas altamente heterocigotas, una de alta frecuencia de pericarpio negro y grano no reventón (A) se cruzó por otra de pericarpio incoloro y grano reventón (B), en la siguiente campaña las semillas híbridas no convencionales (AxB) fueron sembradas, 1200 plantas fueron autofecundadas, en la cosecha, por defectos de la mazorca, pudrición o mal llenado, se eliminaron 400 mazorcas. Finalmente, las 256 mejores mazorcas de grano negro fueron conservadas para los ensayos.
2.3 Características de la parcela experimental. Se utilizó el diseño de Látice Simple 16x16 con 256 líneas S1 (genotipos) y dos repeticiones para cada uno de los tres ensayos. El diseño de lattice es más preciso que el de bloques completos al azar (Cochran, 1941). Los genotipos (tratamientos) fueron asignados a parcelas experimentales de un surco, dentro del surco los golpes estaban distanciados a 35 cm y dos plantas por golpe. La distancia entre surco fue de 80 cm, cada parcela experimental tuvo 12 plantas. La siembra de Huánuco fue realizada el 18 de mayo del 2019 y la cosecha el 26 de octubre del 2019, los experimentos de La Molina se sembraron el 22 de junio de 2019 y se cosecharon el 5 de diciembre del 2019; un ensayo de La molina se fertilizó con 150-0-0 kg/ha de N-P-K, mientras que al otro se aplicó 150-100-100 kg/ha de N-P-K, en cada localidad, todas las líneas fueron sometidas a la misma dosis de fertilización. El manejo agronómico de los tres experimentos -preparación del terreno, control de malezas y control de plagas-, fueron llevados a cabo de acuerdo a las recomendaciones establecidas para un lote comercial de producción de mazorcas de maíz morado.
2.4 Variables a evaluar
Volumen de expansión (VE): La expansión se llevó a cabo siguiendo la metodología de Pordesimo et al. (1990), con modificaciones. Treinta gramos de grano fueron colocados en un sobre de papel kraft N° 10 para la cocción. Al final del período de reventado, se midió el volumen de los mismos en una probeta graduada de 500 ml y se calculó el volumen de expansión de acuerdo a la fórmula: VE = (volumen de granos reventados / peso inicial, 30 g). El método de expansión se ajustó a lo propuesto por Paraginski et al. (2016), la expansión se realizó en un horno microondas (Daewoo, 1000 W, modelo: KOR-32FCB), el tiempo de cocción varió entre 2 y 4 minutos, la expansión se interrumpió cuando hubo un intervalo de tiempo de 5 s entre la expansión de un grano y el siguiente.
Expansión de granos (EG). Se obtuvo con la siguiente fórmula: EG = [(total de granos expandidos) / (total de granos tostados)]*100
Pericarpio negro (PN). Después de la cosecha y en cada tratamiento, se contabilizó el total de mazorcas de pericarpio negro y se expresó en porcentaje para el análisis estadístico.
Rendimiento de grano (RG). Se determinó con la siguiente fórmula: RDG = Pc * 0,971 * ID * Fh * Ff * (10000 m2/1.75 m2), modificado de Sayre et al. (2012), donde: Pc = Peso de mazorcas cosechadas por parcela, 0.971 = Factor de contorno, ID = Índice de desgrane (peso de grano/peso de mazorca), 1.75 m2 = Área de la parcela, Fh = Factor de corrección por humedad, ajustado al 14 %, Ff =Factor de corrección por fallas. Fh =[(100 - % humedad de cosecha) / (100-14)], Ff = [(golpes por parcela - 0,3*fallas por parcela) / (golpes por parcela - fallas por parcela)].
2.5 Análisis de datos. Se realizó utilizando el software estadístico R versión 4.0.4 (2021-02-15) de la R Foundation for Statistical Computing, y los paquetes agricolae versión 1.3-3 (Felipe de Mendiburu, 2020), ggplot2 versión 3.3.3. (Wickham, 2016) y PerformanceAnalytics versión 2.0.4. (Peterson et al., 2018).
3. Resultados y discusión
3.1 Modo de segregación de los caracteres de valor
Los caracteres volumen de expansión, expansión de granos, pericarpio negro y rendimiento de grano, evaluados en las 256 líneas S1, parecen tener herencia del tipo cuantitativa o poligénica debido a que sus histogramas de frecuencias mostraron variación continua en forma de campana (Figura 1), modo de segregación similar a lo propuesto por Nilsson-Ehle (1909), Klug et al. (2017) y Zhang et al. (2018) para estas características, sin embargo, en el histograma del pericarpio negro, además de la distribución continua, se puede notar la presencia de tres modas, la primera alrededor de 57,5%, la segunda alrededor de 67,5% y la última alrededor de 80,0%, lo cual es indicativo de la presencia de genes de efectos mayores que están segregando. Las 256 líneas S1 evaluadas son segregantes del cruce de dos poblaciones, una con 0 de VE y otra con 25,7 ml/g de VE, en esta generación S1 (primera autofecundación de plantas híbridas y equivalente a la F2 de plantas autógamas) se aprecia que las líneas presentaron valores bajos de volumen de expansión, cuyo promedio fue de 2,97 ml/g, valor muy inferior al promedio de las poblaciones parentales (12,85 ml/g), esta distribución continua sesgada hacia la izquierda pondría en evidencia la existencia de cierta dominancia de genes con valores bajos de VE, estos resultados concuerdan con los hallados por Crumbaker et al. (1949); sin embargo, el hecho de tener una distribución en forma de campana pone en evidencia de que los efectos genéticos aditivos son más importantes, como lo señalan Pereira & Amaral Júnior (2001), Arnhold et al. (2010) y Coan et al. (2019). En relación a la expansión de granos, este porcentaje varió de 2,1 a 62,3, esta característica también mostró variación continua, considerando que los progenitores de los híbridos provienen del cruce de poblaciones con 0% y 94% de granos expandidos, es evidente que -en esta generación S1 que equivale a la F2-, hay cierta dominancia de genes con valores bajos ya que la mayoría de los valores están por debajo de la media esperada de 47%, en el caso de ausencia de dominancia; sin embargo, los efectos de genes aditivos también son importantes. En la producción de antocianina (Tamay, 2021) y el rendimiento de grano (Valera, 2019) el efecto de los pisos altitudinales no es tan importante. El color negro del pericarpio, no se presentó como una característica de herencia cualitativa, sino más bien, se asemejó a la de un carácter cuantitativo, como lo evidenciaron Chatham & Juvik (2021) y Sa et al. (2020). También tuvo distribución continua (Figura 1) y fluctuó entre líneas, de 25,4% a 98,3% de mazorcas con granos negros. La segregación fue alta, por lo que se pudo apreciar colores variables en pocas mazorcas (Figura 2). A diferencia de lo reportado por Ford (2000), en donde fue posible encontrar proporciones clásicas de 3/4-1/4; 12/16-3/16-1/16, etc., en esta investigación el uso de progenitores de los híbridos altamente heterocigotos impidió que se obtengan los mismos resultados. El rendimiento de grano fue otra característica de distribución continua, los valores fenotípicos se presentaron dentro de un rango de 0,9 a 5,7 t/ha, considerando que los rendimientos de los progenitores son de 1,5 t/ha y 6 t/ha, se esperaría un rendimiento promedio alrededor de 3,75 t/ha, sin embargo, en promedio, las líneas rindieron 2,3 t/ha, muy por debajo de lo esperado, esto probablemente se debe a la pérdida de vigor como fue señalado por Jones (1918), y a que estas genotipos tienen una generación de autofecundación el cual aumentó la endogamia en un 50%. La distribución continua de los fenotipos (Figura 1) es indicativo de que este carácter también es de herencia poligénica.
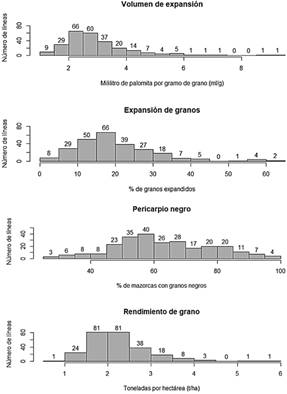
Figura 1 Histograma de frecuencias de cuatro caracteres a partir de los datos de 256 líneas segregantes de maíz morado reventón.

3.2 Selección de líneas S1 de maíz morado reventón
En las variedades mejoradas de maíz reventón, el carácter de valor es el volumen de expansión, debido que el vendedor de maíz compra en magnitud de peso (granos) para vender en volumen (palomitas); sin embargo, en variedades de maíz morado reventón, además, se tiene que considerar que el pericarpio sea pigmentado (negro). En la selección de caracteres cuantitativos es de vital importancia seleccionar un número grande de individuos, líneas en este caso, para aumentar la probabilidad de obtener alelos favorables que controlen las características de interés, que, frecuentemente, se encuentran dispersos en diferentes genotipos. Previo a la selección simultánea por VE y PN, se realizó el análisis de correlación entre las cuatro características (Figura 3), cuyo resultado evidenció una correlación negativa entre los caracteres de valor de la futura variedad, VE y PN; asimismo, considerando que en promedio del total de líneas, el valor del VE fue de 2,97 ml/g, valor muy por debajo del grano reventón amarillo importado que alcanza los 30 ml/g de Oliveira et al. (2020), y el del PN de 62,8%, valor cercano a lo deseado, basado en la selección simultanea e intencional para múltiples caracteres como lo explican Bos & Caligari (2007), se propuso la siguiente fórmula para estimar el índice de selección [IS=(0,9*VE)+(0,1*PN)], en la que el peso asignado a VE fue de 0,9 y al PN de 0,1, valores del IS que permitieron identificar a las líneas sobresalientes (Tabla 1), con este criterio, se logró seleccionar 80 líneas que representan una presión de selección del 31,25%. Si bien, la selección se hizo en base a los atributos VE y PN, también es posible estimar el diferencial de selección (DS) para los caracteres que no se seleccionaron, como la EG y el RG (parte final de la Tabla 1), en ella se puede notar que el DS fue positivo para VE, PN, y EG; sin embargo, no lo fue para RG, aparentemente, cuando se mejora por calidad se disminuye el rendimiento de grano, tal como lo reportó Gimenez (2017).
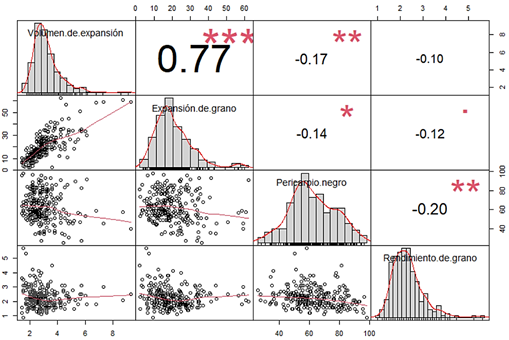
Figura 3 Coeficiente de correlación entre VE, EG, PN y RG a partir de los datos de 256 líneas segregantes de maíz morado reventón.
Tabla 1 Relación de líneas sobresalientes seleccionadas por volumen de expansión y pericarpio negro
| N° de orden | Línea S1 seleccionada | Volumen de expansión (ml/g) | Expansión de granos (%) | Pericarpio negro (%) | Rendimiento de grano (t/ha) | Índice de selección |
| 1 | 183 | 6,1 | 29,9 | 91,7 | 1,8 | 14,6 |
| 2 | 136 | 5,3 | 24,6 | 89,5 | 2,1 | 13,7 |
| 3 | 79 | 8,7 | 60,7 | 57,6 | 2,1 | 13,6 |
| 4 | 122 | 5,7 | 35,2 | 82,7 | 3,1 | 13,4 |
| 5 | 43 | 4,2 | 62,3 | 90,8 | 2,0 | 12,9 |
| 6 | 169 | 4,7 | 10,9 | 86,2 | 2,4 | 12,8 |
| 7 | 144 | 3,5 | 41,5 | 92,8 | 2,0 | 12,4 |
| 8 | 131 | 4,5 | 20,5 | 83,2 | 1,5 | 12,4 |
| 9 | 152 | 9,3 | 59,0 | 39,6 | 2,5 | 12,3 |
| 10 | 237 | 3,9 | 28,2 | 86,3 | 1,6 | 12,2 |
| 11 | 86 | 3,1 | 34,7 | 93,4 | 2,0 | 12,1 |
| 12 | 71 | 2,8 | 28,1 | 95,8 | 1,5 | 12,1 |
| 13 | 109 | 6,8 | 58,6 | 59,0 | 1,8 | 12,0 |
| 14 | 64 | 3,6 | 19,6 | 85,6 | 2,8 | 11,8 |
| 15 | 41 | 3,2 | 13,4 | 87,1 | 1,5 | 11,6 |
| 16 | 30 | 2,2 | 15,5 | 96,4 | 2,3 | 11,6 |
| 17 | 16 | 2,8 | 24,0 | 90,4 | 2,1 | 11,6 |
| 18 | 249 | 2,7 | 17,3 | 90,5 | 1,5 | 11,5 |
| 19 | 141 | 2,6 | 24,8 | 91,5 | 1,3 | 11,5 |
| 20 | 234 | 3,6 | 29,5 | 82,5 | 1,7 | 11,5 |
| 21 | 35 | 4,2 | 35,6 | 76,5 | 1,7 | 11,4 |
| 22 | 27 | 3,6 | 2,1 | 81,5 | 4,5 | 11,4 |
| 23 | 247 | 1,5 | 9,5 | 98,3 | 0,9 | 11,2 |
| 24 | 38 | 2,6 | 16,2 | 88,5 | 1,8 | 11,2 |
| 25 | 231 | 2,9 | 21,2 | 85,5 | 1,4 | 11,2 |
| 26 | 51 | 7,3 | 59,0 | 45,7 | 3,0 | 11,2 |
| 27 | 13 | 3,2 | 27,6 | 82,2 | 1,8 | 11,1 |
| 28 | 238 | 3,3 | 33,8 | 81,3 | 1,6 | 11,1 |
| 29 | 130 | 4,2 | 34,6 | 72,3 | 1,9 | 11,0 |
| 30 | 5 | 3,3 | 33,1 | 79,9 | 3,0 | 11,0 |
| 31 | 178 | 2,7 | 19,1 | 85,3 | 2,4 | 11,0 |
| 32 | 111 | 3,2 | 27,5 | 80,6 | 1,7 | 10,9 |
| 33 | 250 | 1,4 | 6,4 | 96,2 | 1,2 | 10,9 |
| 34 | 158 | 2,9 | 27,0 | 83,0 | 1,8 | 10,9 |
| 35 | 134 | 4,0 | 40,6 | 72,7 | 2,6 | 10,9 |
| 36 | 77 | 2,2 | 12,1 | 88,9 | 1,9 | 10,8 |
| 37 | 57 | 3,5 | 24,2 | 76,4 | 2,2 | 10,8 |
| 38 | 59 | 3,2 | 17,3 | 78,9 | 1,3 | 10,8 |
| 39 | 242 | 4,1 | 12,1 | 70,7 | 1,9 | 10,8 |
| 40 | 46 | 3,2 | 20,1 | 78,6 | 2,6 | 10,7 |
| 41 | 121 | 3,1 | 12,5 | 79,2 | 1,6 | 10,7 |
| 42 | 48 | 3,9 | 23,7 | 71,7 | 1,6 | 10,7 |
| 43 | 162 | 2,9 | 13,5 | 81,0 | 1,3 | 10,7 |
| 44 | 3 | 6,0 | 33,8 | 53,3 | 2,4 | 10,7 |
| 45 | 65 | 5,5 | 26,5 | 56,8 | 1,8 | 10,7 |
| 46 | 125 | 2,2 | 3,5 | 86,3 | 2,4 | 10,6 |
| 47 | 101 | 2,5 | 25,0 | 83,6 | 2,6 | 10,6 |
| 48 | 56 | 5,6 | 34,1 | 55,4 | 2,2 | 10,6 |
| 49 | 17 | 2,9 | 28,9 | 79,4 | 1,4 | 10,6 |
| 50 | 126 | 3,6 | 18,1 | 73,0 | 1,9 | 10,6 |
| 51 | 115 | 2,2 | 15,8 | 85,4 | 1,7 | 10,6 |
| 52 | 68 | 2,4 | 17,8 | 83,4 | 2,1 | 10,5 |
| 53 | 4 | 2,5 | 17,4 | 82,3 | 2,0 | 10,5 |
| 54 | 117 | 3,7 | 34,1 | 70,8 | 2,5 | 10,4 |
| 55 | 147 | 2,2 | 11,9 | 84,3 | 2,8 | 10,4 |
| 56 | 54 | 2,7 | 23,7 | 80,0 | 2,8 | 10,4 |
| 57 | 97 | 2,7 | 14,5 | 79,0 | 1,8 | 10,3 |
| 58 | 189 | 2,1 | 14,6 | 84,4 | 2,3 | 10,3 |
| 59 | 123 | 3,2 | 22,5 | 74,3 | 1,7 | 10,3 |
| 60 | 244 | 2,8 | 25,0 | 77,8 | 4,0 | 10,3 |
| 61 | 73 | 2,2 | 14,3 | 82,3 | 2,2 | 10,2 |
| 62 | 11 | 2,7 | 17,4 | 77,6 | 3,8 | 10,2 |
| 63 | 218 | 3,2 | 19,0 | 73,5 | 1,6 | 10,2 |
| 64 | 167 | 3,9 | 27,6 | 66,4 | 1,6 | 10,2 |
| 65 | 124 | 2,3 | 15,9 | 80,8 | 1,7 | 10,2 |
| 66 | 69 | 2,4 | 7,6 | 78,5 | 3,7 | 10,0 |
| 67 | 214 | 2,2 | 11,7 | 80,7 | 2,2 | 10,0 |
| 68 | 177 | 4,4 | 24,0 | 60,2 | 2,7 | 10,0 |
| 69 | 161 | 3,7 | 24,2 | 66,9 | 1,8 | 10,0 |
| 70 | 212 | 2,0 | 10,7 | 82,0 | 1,6 | 10,0 |
| 71 | 254 | 2,1 | 20,7 | 80,9 | 2,3 | 10,0 |
| 72 | 114 | 1,8 | 9,0 | 83,8 | 3,0 | 10,0 |
| 73 | 90 | 2,7 | 30,8 | 75,3 | 1,6 | 9,9 |
| 74 | 216 | 2,3 | 16,9 | 78,1 | 1,5 | 9,9 |
| 75 | 128 | 3,4 | 37,2 | 68,2 | 1,7 | 9,9 |
| 76 | 32 | 4,6 | 56,6 | 56,9 | 2,4 | 9,9 |
| 77 | 160 | 3,1 | 16,5 | 70,1 | 1,7 | 9,8 |
| 78 | 112 | 4,8 | 32,9 | 54,3 | 2,3 | 9,8 |
| 79 | 2 | 5,6 | 50,8 | 47,1 | 2,5 | 9,8 |
| 80 | 139 | 3,4 | 19,5 | 66,6 | 2,5 | 9,8 |
| Promedio de 80 líneas selectas | 3,5 | 25,0 | 77,8 | 2,1 | ||
| Promedio de 256 líneas | 3,0 | 20,0 | 62,8 | 2,3 | ||
| Diferencial de selección | 0,5 | 5,0 | 15,0 | -0,2 | ||
| Rango de 256 líneas | 1,4 - 9,3 | 2,1 - 62,3 | 25,4 - 98,3 | 0,9 - 5,7 | ||
4. Conclusiones
Los caracteres: volumen de expansión (VE), expansión de granos (EG), pericarpio negro (PN) y rendimiento de grano (RG), evaluados en 256 líneas S1 obtenidas por autofecundación a partir de híbridos no convencionales, mostraron variación continua cuyos histogramas indican que la acción de alelos aditivos son los más importantes. Además de la distribución continua, en los caracteres VE, EG y RG, se observó cierta dominancia de genes con valores bajos, mientras que para PN se notó la presencia de tres modas, los cuales indicarían la presencia de genes de efectos mayores. Así mismo, considerando el modo de distribución de los caracteres de valor VE y PN, se estableció un índice de selección que permitió identificar a 80 líneas sobresalientes que representan una presión de selección de 31,25%. Con las líneas seleccionadas, se espera: 1) llevarlas a homocigosis total a través del cultivo de esporas haploides o del uso de la tecnología de dobles haploides, ya que en S1 aún es necesario depurar alelos no deseados, 2) formar híbridos simples a partir de líneas 100 % homocigotas, y 3) recombinarlas para obtener una población de libre polinización de amplia base genética, con mayor frecuencia de genes favorables y con mejores valores para VE y PN, a partir del cual, iniciar un nuevo ciclo de selección.

