











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
El bosque alto andino se caracteriza como un ecosistema que está ubicado entre los 2400 y 5400msnm aproximadamente, el cual forma parte del perímetro inferior del ecosistema de páramo (Blancas et al., 2020; Ortiz et al., 2021; Cardona Iglesias et al., 2021). Está compuesto por una gran diversidad de plantas nativas con rasgos morfológicos que confluyen en este hábitat y regulan los servicios ambientales que ahí se ofrecen; agua, oxigeno, madera, leña, flora y fauna (Bacca A. y Burbano M., 2018; Bacca et al., 2020), además alberga diversidad importante, debido a su alta riqueza específica y porcentaje de endemismo (Obregón-La Rosa et al., 2021; Quispe Rojas y Elias Nuñez, 2020).
Pese a esto, el manejo de este bioma de alta fragilidad no es el mejor, los ecosistemas se enfrentan con retos difíciles de superar, tanto en desarrollo sustentable como en el incremento de nuevas áreas con la misma aptitud. Estos retos están relacionados con el cambio climático, el manejo ambiental y la agricultura convencional de montaña, así como la repercusión de la globalización socio- económica y cultural; los cuales tienen a este ecosistema al borde de la desaparición (Haller y Branca, 2020).
Los sistemas productivos que comúnmente prevalecen, como la ganadería intensiva de leche y el cultivo agrícola de papa ejercen alta presión y fragmentación de los sistemas naturales. El uso intensivo de sistemas agrícolas ha traído consigo la perdida de diversidad ecológica, bajos rendimientos, erosión y ampliación de la frontera agrícola en zonas estratégicas o de recarga hídrica (Ruiz et al., 2018). Se dice que los sistemas productivos intensivos son los responsables a nivel mundial del 80% de la deforestación (Wilson et al., 2020). A causa de la expansión de la frontera agrícola y la ganadería intensiva se establece que traerá consigo la perdida de al menos el 25 por ciento de la biodiversidad en los próximos años, además las fluctuaciones en el clima afectan considerablemente su biodiversidad (Reyes-Palomino y Cano Ccoa, 2022). El desconocimiento de biotecnologías apropiadas para estos ecosistemas inducirá a sistemas productivos económicamente poco rentables, ambientalmente insostenibles y socialmente inadecuados (Lanz et al., 2018).
Referente al contexto socio cultural y económico, los campesinos de la región haciendo uso de prácticas convencionales propiciadas por la revolución verde ha conllevado que amplíen sus áreas productoras hacia nuevas zonas de ausente vocación agropecuaria, como páramos y subpáramos, motivados por la necesidad de encontrar suelos más productivos para satisfacer las necesidades básicas de alimento, vivienda y económicas, que aun así, no son los mejores ya que los niveles de desnutrición y pobreza son considerables (Candelaria et al., 2020), provocando a su vez un desentendimiento del uso sostenible de los recursos naturales de alta montaña.
Bajo estas circunstancias se crea la necesidad de buscar estrategias que permitan la recuperación de manera integral este importante bioma. La identificación, caracterización de rasgos morfológicos y domesticación de especies nativas son alternativas, que pueden contribuir a la conservación y restauración de zonas degradadas mediante la regulación del recurso hídrico, enriquecimiento de la biodiversidad y la interconectividad de paisajes (FAO 2019). Con este tipo de estudios, la flora nativa a través de sistemas agroecológicos como los Sistemas agroforestales-SAF pueden tener un uso productivo sin colocar en riesgo la estructura ecológica y la biodiversidad de la región, permitiendo a su vez obtener una estrategia eficiente donde se involucra el ámbito socio cultural, ambiental y económico cuyo objetivo primordial es contribuir al desarrollo sostenible de los sistemas productivos prevalecientes de la región (Rodríguez Espinosa et al., 2020).
En este sentido, estudiar los rasgos morfológicos de especies potenciales para este fin se hace necesario, ya que se requiere ampliar el abanico o la disponibilidad de especies útiles para arreglos SAF que sean propios de este ecosistema. Los rasgos morfológicos son expresiones de cada una de las especies, que determinan el crecimiento, supervivencia y reproducción, además pueden actuar como indicadores ecológicos ante cambios ambientales o antrópicos (Romero-Saritama y Pérez- Ruiz, 2016). Según Moreno et al. (2015), los estudios de caracterización morfológica, en particular los atributos de hojas, se pueden basar en caracteres cualitativos o cuantitativos que determinan el uso y disposición de los recursos del medio. Además, aporta información en la taxonomía, detección de plagas y enfermedades, protección contra pesticidas, alimentación, medicina, industria y medio ambiente (Cervantes et al., 2017). Así como también en la identificación de individuos que pueden ser útiles para fuentes semilleras en programas de restauración ecológica y sistemas agroforestales (Chao et al., 2019).
En la actualidad se ha analizado rasgos foliares en especies arbóreas nativas para diferentes zonas agroecológicas, sin embargo, para las condiciones ambientales del bosque alto andino los estudios son limitados (Castañeda-Garzón et al., 2021) y la poca información que se tiene no es actual. Por lo tanto, se hace relevante investigar a especies nativas como Cleome arborea Kunth; Weinmannia mariquitae Szyszyl; Viburnum triphyllum Benth y Leandra subseriata (Naudin).
Las investigaciones realizadas por Bacca y Burbano (2018); Bacca et al. (2020); Bernal y Celis (2020); Melissa y Torres (2016), establecen que las especies antes mencionadas favorecen a servicios ecosistémicos como: regulación del recurso hídrico, captura de carbono y fuente de alimento para animales silvestres, por lo tanto, brindar información sobre su caracterización a través de descriptores morfológicos de hojas se hace necesaria para procesos de identificación, colecta, propagación y domesticación que permitirá recuperar las condiciones propias del ecosistema que habitan.
En la limitada investigación que se encontró, Jáuregui y Ruiz-Zapata (2011), mencionan que el género Cleome es uno de los más números con aproximadamente 200-250 taxones. Los trabajos que hasta el momento se han generado en relación con este estudio están encaminados a la evaluación de la estructura de lámina foliar como mecanismo fotosintético (Marshall et al., 2007; Muhaidat et al., 2007; Voznesenskaya et al. 2007). Aún es necesario que se indague sobre este importante ecosistema junto con la flora existente, conocer en detalle su dinámica de crecimiento y su morfología permitirá tomar decisiones más acertadas para su conservación y uso sostenible mediante sistemas agroecológicos.
Bajo este contexto dentro del marco del macroproyecto denominado “Estudio fenológico de la flora silvestre nativa con potencial para procesos de restauración ecológica en bosque alto andino del municipio de Pasto, departamento de Nariño” el objetivo de la investigación fue determinar los patrones morfológicos foliares que caracterizan a C. arborea; W. mariquitae; V. triphyllum y L. Subseriata en el bosque Alto Andino en la Granja Experimental Botana de la Universidad de Nariño, que contribuya al estudio y domesticación de las mismas, para involucrarlas en futuras investigaciones en sistemas agroecológicos como SAF que permitan la recuperación y resiliencia de los ecosistemas de alta montaña.
Materiales y métodos
Área de estudio
Este trabajo se desarrolló en la zona de bosque alto andino de la Granja Experimental Botana de la Universidad de Nariño. Corregimiento de Catambuco, Municipio de Pasto, departamento de Nariño, cuyas coordenadas son: 01°09’29.37” N y 77°16’33.24” O, a una altura de 2820msnm, con temperatura promedio de 13°C, el nivel de precipitación aproximadamente es de 967mm anual y 75% de humedad relativa (Bacca et al., 2020). Área perteneciente a la zona de vida bosque húmedo montano bajo (bh - MB) (Zapata-Molina et al., 2020).
Caracterización morfológica
De acuerdo con la metodología propuesta por Cervantes et al. (2017), se tomaron en total 10 individuos por cada especie en los cuales se evaluaron 15 descriptores de hojas (Tabla 1). Se recolectaron dos hojas compuestas para las especies de C. arborea y W. mariquitae en ellas se muestrearon cinco foliolos centrales ubicados a los dos costados de la hoja, mientras que para V. triphyllum y L. subseriata se utilizaron de 10-20 hojas simples para cada especie dependiendo de las condiciones de acceso y fitosanitarias.
Las muestras presentaron características como: hojas maduras, completas y sin problemas fitosanitarios (figura 1), ubicadas en los cuatro puntos cardinales de las ramas de la parte media y periférica de la copa (Castañeda-Garzón et al., 2021). Las hojas fueron colocadas en bolsas de papel previamente etiquetadas con la especie y número de individuo. Con el propósito de evitar la deshidratación, los datos morfométricas cualitativos y cuantitativos se registraron inmediatamente después de la colecta, para estos últimos se utilizó un calibrador digital de precisión 0,1mm marca Truper (Truper Inc., Colombia) (Castañeda-Garzón et al., 2021) y la estimación cromática de la lámina foliar se realizó mediante inspección visual directa con la carta de colores de Munsell, bajo condiciones de iluminación estandarizadas (Kehlenbeck et al., 2015).

Figura 1 Medidas morfométricas de hojas. A; Weinmannia mariquitae Szyszyl, B; Viburnum triphyllum Benth, C; Leandra subseriata Naudin, D; Cleome arborea Kunth.
Tabla 1 Descriptores cuantitativos y cualitativos de hoja
| No | Descriptor | Variable | Abreviatura | No | Descriptor | Variable | Abreviatura |
|---|---|---|---|---|---|---|---|
| 1 | Longitud pecíolo principal | Cuantitativa | LPEC ppal | 10 | Pubescencia envés | Cualitativa | PUBENV |
| 2 | Grosor pecíolo principal | Cuantitativa | GPEC | 11 | Color hoja haz | Cualitativa | CHHAZ |
| 3 | Largo hoja | Cuantitativa | LH | 12 | Color hoja envés | Cualitativa | CHENV |
| 4 | Ancho hoja | Cuantitativa | ANH | 13 | Ondulación Lámina Foliar | Cualitativa | OH |
| 5 | Forma hoja | Cualitativa | FH | 14 | Venación haz | Cualitativa | VHAZ |
| 6 | Borde hoja | Cualitativa | BORH | 15 | Venación envés | Cualitativa | VENV |
| 7 | Ápice hoja | Cualitativa | APH | ||||
| 8 | Base hoja | Cualitativa | BH | ||||
| 9 | Pubescencia haz | Cualitativa | PUBHAZ |
Fuente: Elaboración propia.
Análisis estadístico
El registro de los datos se realizó en una matriz de Excel discriminando la especie. La aplicación de las pruebas estadísticas se realizó a través del programa InfoStat® 2020. Los datos morfométricos cuantitativos se sometieron al análisis de componentes principales (ACP) mientras que a los datos cualitativos se les aplicó un análisis de frecuencias con el fin de categorizarlos para posteriormente realizar el análisis de correspondencias múltiples (ACM) (Castañeda-Garzón et al., 2021; Meneses, 2019).
Resultados
En la tabla 2 se describe de manera general las cuatro especies evaluadas, información que parte de los datos de frecuencia de descriptores cualitativos estimados.
Tabla 2 Caracterización de descriptores predominantes de las especies C. arborea W. mariquitae V. triphyllum y L. subseriata
| C. arborea | W. mariquitae | V. triphyllum | L. subseriata | |||||
|---|---|---|---|---|---|---|---|---|
| Descriptor | Categoría | % | Categoría | % | Categoría | % | categoría | % |
| FH | Palmatisecta | 79 | Elíptica | 100 | Elíptica | 70 | Ovada | 56 |
| BORH | Entero | 45 | Serrulado | 100 | Entero | 100 | Aserrado | 100 |
| APH | Acuminado | 81 | Redondeado | 83 | Apiculado | 100 | Agudo | 86 |
| BH | Cuneada | 77 | Redondeada | 73 | Redondeada | 53 | Obtuso | 64 |
| PUBHAZ | Presente | 100 | Presente | 100 | Presente | 51 | Presente | 100 |
| PUBENV | Presente | 100 | Presente | 100 | Ausente | 100 | Presente | 100 |
| CHHAZ | 7,5GY 4/6 | 22 | 7,5GY 4/6 | 64 | 7,5GY 4/4 | 32 | 7,5GY 5/8 | 37 |
| CHMENV | 5GY 7/10 | 30 | 5GY 6/4 | 16 | 7.5GY 5/4 | 22 | 5GY 7/10 | 38 |
| OH | Ondulada | 100 | Plana | 99 | Plana | 100 | Ondulada | 100 |
| VHAZ | Hundida | 100 | Hundida | 100 | Hundida | 82 | Intermedia | 58 |
| VVEN | Hundida | 100 | Alzada | 100 | Alzada | 91 | Alzada | 63 |
Fuente: Elaboración propia.
De igual manera, como complemento de la información descriptiva presentada anteriormente, en la tabla 3 se observa las medidas resumen de los registros encontrados para C. arborea W. mariquitae V. triphyllum y L. subseriata.
Tabla 3 Medidas morfométricas de las variables cuantitativas para las especies C. arborea W. mariquitae V. triphyllum y L. subseriata
| Especie | C. arborea | W. mariquitae | V. triphyllum | L. subseriata | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Descriptor | LI (mm) | x̄ (mm) | LS (mm) | SD (mm) | LI (mm) | x̄ (mm) | LS (mm) | SD (mm) | LI (mm) | x̄ (mm) | LS (mm) | SD (mm) | LI (mm) | x̄ (mm) | LS (mm) | SD (mm) |
| LPECppal | 95,7 | 147,5A | 199,2 | 40.41 | 8,8 | 20,7B | 45,8 | 6,7 | 3,8 | 7,9C | 13,0 | 1,8 | 3,2 | 5,9C | 8,6 | 1,3 |
| GPEC | 2,8 | 6,7A | 10,6 | 2,6 | 1,0 | 1,7B | 2,5 | 0,3 | 0,9 | 1,6BC | 2,8 | 0,4 | 0,2 | 0,8C | 1,5 | 0,3 |
| LH | 34,9 | 118,6A | 202,4 | 34,1 | 42,9 | 91,7B | 140,8 | 19,8 | 47,0 | 76,1C | 107,6 | 14,5 | 8,1 | 14,0D | 19,9 | 2,7 |
| ANH | 16,6 | 32,9A | 49,2 | 32,5 | 22,0 | 38,8B | 61,7 | 8,2 | 18,4 | 34,9B | 53,5 | 8,1 | 4,0 | 9,2C | 14,3 | 2,0 |
Letras diferentes difieren significativamente. Tukey; Alfa=0,05. LI; límite inferior, X; media, LS; límite superior, SD; desviación estándar.
Fuente: Elaboración propia.
El ACP para C. arborea permitió agrupar las variables en dos componentes principales que explicaron el 69% (figura 2A) de la varianza total de la muestra. El PC1 explicó el 37% de la varianza total. En su medida, las cuatro variables aportaron a la conformación de dicho factor; ANH (r=0,73), LH (r=0,70), GEPC (r=-0,54) y LPEC ppal (-0,12), sin embargo, si tomamos en cuenta el coeficiente de correlación podemos observar que ANH y LH fueron las más importantes. El segundo componente explicó el 33% de la variabilidad el cual se encuentra influenciado por GEPC (r=0,56), LH (r=0,41), LPECppal (r=0,87) y ANH (r=0,28), en el cual la variable LPEC ppal tuvo mayor relevancia. Además, teniendo en cuenta el ángulo de los vectores se puede establecer que existe una estrecha correlación entre ANH-LH (P=0,0008) con el 60%, mientras que si comparamos ANH con LPEC ppal no presenta correspondencia.
De acuerdo con ACP para L. subseriata agrupó las variables en dos componentes principales que explicaron el 83% de la variación total. El PC1 explicó el 61% de la variación. Las variables evaluadas aportaron de manera diferencial sobre este factor; ANH (r=0,87), LH (r=0,87), GEPC (r=-0,82) y LPEC ppal (r=0,52). Sin embargo, si consideramos la información presentada en la figura 2B y el coeficiente de correlación mencionado anteriormente podemos establecer que LH y ANH son las dos primeras variables morfológicas que más se destacan para esta especie. El segundo componente explico el 22% de la variabilidad a diferencia de LPEC ppal (r=0,84), los valores de correlación de las variables son bajos con respecto al componente uno; GEPC (r=-0,35), ANH (r=-0,23) y LH (r=0,07). Por otra parte, Los individuos formaron dos grupos; las variables que definieron el primer grupo fueron LPEC ppal-LH (P=<0,0001) mientras que el segundo grupo estuvo conformado por GPEC-ANH (P=<0,0001).
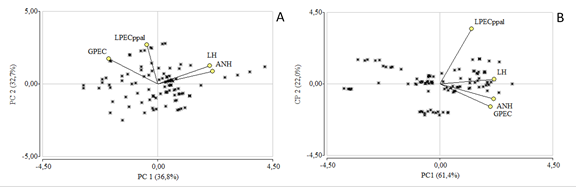
Figura 2 Análisis de componentes principales de los rasgos morfológicos foliares. A. C. arborea . Correlación cofenética=1,00. B. L. subseriata. Correlación cofenética=1,00.
Para las variables cualitativas de C. arborea, la contribución de los cinco primeros ejes explica un porcentaje de 51,79% de la variabilidad total. El primer eje explicó el 25,58%, el segundo; 10,43%, el tercero 5,82%, el cuarto; 5,36% y el quinto 4,60%. El 50% de la muestra presentó una correlación positiva con la variable APH; 10_11, CHHAZ; 8, CHENV; 11 y correlación inversa con CHHAZ; 4_8_11_9 y CHENV; 7-8. El 50% restante se correlaciono con las variables CHHAZ; 1_10_4_11 y CHENV; 4_9_1_3_8-11. Si en la misma tabla se contempla los aportes realizados por cada variable en los diferentes componentes se puede observar que FH , APH, BH, CHHAZ, CHENV tuvieron los valores más altos.
Para las variables cualitativas de L. subseriata la contribución de los cinco primeros factores explica un porcentaje de 40,58% de la variabilidad total. El primer factor explicó el 9,87%, el segundo factor 8,41%, el tercer factor 8,08%, el cuarto factor 7,18% y el quinto factor 7,03%. Se forman tres grupos; el primero presenta correlación con las variables BH 2, CHHAZ 5_4 y CHENV 8. El segundo grupo se correlacionó con BH_4, VHAZ 2_1, FH 4_1, VENV 2_1; BORH 1; PUBHAZ 1, PUBENV 1, CHENV 10_7, CHEHAZ 1_7_2. El tercer grupo presentó asociación con las variables CHEHAZ 3_8, CHENV 1_9, BH 1_3. Si se contempla los aportes realizados por cada variable en los diferentes componentes, se establece que BH, CHHAZ, CHENV, APH fueron los descriptores más importantes; información relevante para análisis descriptivo de esta especie sobre este ecosistema.
Para W. mariquitae en el ACP se agruparon las variables en dos componentes principales que explicaron el 89% de la varianza. El PC1 explicó el 64% de la varianza total. Las variables con mayor coeficiente de correlación en orden decreciente son; ANH (r=0,95), LH (r=0,94), LPECppal (r=0,88) y GPEC (r=0,06). Además, observando el ángulo de los vectores (figura 3A) se puede apreciar una estrecha relación entre estas tres primeras variables. En este sentido, serían los descriptores morfológicos que caracterizan a esta especie para su identificación. El segundo componente explicó el 25% de la variabilidad, en el cual a diferencia de GPEC (r=0,87), los valores de correlación son bajos; LPECppal (r=0,08), ANH (r=-0,07) y ANH (r=-0,06). En el gráfico se identifica un grupo que correlaciona todas las variables de la siguiente manera: ANH-LH (P=<0,0001), LPECppal-LH (P=<0,0001), LPECppal-ANH (P=<0,0001).
En este sentido el ACP de V. triphyllum permitió agrupar las variables en dos componentes principales que explicaron el 88% de la varianza total de la población (figura 3B). El PC1 manifestó el 69% de la varianza total. Las variables con mayor coeficiente de correlación y que más aportaron a la conformación de dicho factor fueron ANH (r=0,88), LH (r=0,86), GEPC (r=-0,80) y LPEC ppal (r=0,78). El segundo componente explicó el 20% de la variabilidad, el cual se encuentra influenciado por LPEC ppal (r=0,48), GEPC (r=0,44), LH (r=-0,44) y ANH (r=-0,40). Se definió un grupo con todas las variables; LPEC ppal, ANH, LH y GPEC (P=<0,0001).
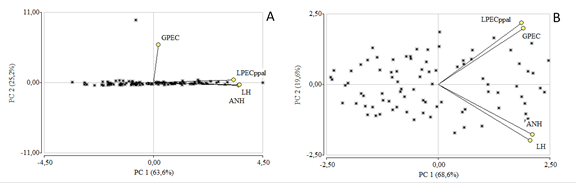
Figura 3 Análisis de componentes principales de los rasgos morfológicos foliares. A. W. mariquitae. Correlación cofenética=0,93. B. V. triphyllum. Correlación cofenética=1,00.
Para las variables cualitativas de W. mariquitae la contribución de los cinco primeros factores explica un porcentaje de 35,34% de la variabilidad total. El primer factor explico el 9,4%, el segundo factor 7,8%, el tercer factor 6,66%, el cuarto factor 6,58% y el quinto factor 5,90%. Se forman tres grupos; el primero correlaciona las categorias BH_1, CHHENV 1_5_8_10_12_15, CHHAZ_5 y APH_ 2. El segundo grupo está conformado por CHHAZ_1, CHENV_1_4_6_7_11_14, PUBHAZ_1, VHAZ_1, OH_1, APH_1 y BH_2. Y el tercer grupo lo conforman CHENV_2_9_13, CHHAZ_2_6_14 y BH_3.
Para las variables cualitativas de V. triphyllum la contribución de los cinco primeros factores explicó un porcentaje de 29,54% de la variabilidad total. El primer factor explico el 6,84%. El segundo factor 6,24%. El tercer factor 5,88%. El cuarto factor 5,54% y el quinto factor 5,03 %. Se correlacionaron las siguientes categorías PUBHAZ_2, VHAZ_2, CHHAZ 13_16_4_ 12, CHENV 1_2_3_5_8_10, FH_2, BH_3. Si se contempla los aportes realizados por cada variable en los diferentes componentes se establece que BH, CHHAZ, CHENV fueron los descriptores de mayor valor.
Discusión
La información acerca de los descriptores cualitativos y cuantitativos, permite conocer los rasgos morfológicos y taxonómicos que caracterizan a C. arborea W. mariquitae V. triphyllum y L. subseriata. Teniendo en cuenta su relevancia descrita en el inicio del documento, esta información coadyuva a procesos de identificación y recolección del material vegetal en campo, que pueden contribuir a programas de restauración, enriquecimiento vegetal y modelos agroecológicos adaptados a la región, en zonas perturbadas por acciones antrópicas o naturales, información que en la actualidad es escasa.
De la misma manera lo menciona Melissa y Torres (2016) quienes indican que la investigación científica sobre caracterización de vegetación en este ecosistema aún es incipiente, se hace necesario hacer estudios que describa a nivel de detalle la cobertura vegetal de este ecosistema de alta fragilidad, el cual según Roa-García y Torres-González (2021), se ve afectado drásticamente por disturbios de tipo agrícola y pecuario. Sumado a esto la información en revistas científicas que concentren sus esfuerzos de investigación en ecosistemas de montaña son limitadas, exceptuando la revista de investigaciones altoandinas de Perú quién en sus últimos volúmenes ha conllevado a brindar conocimientos de alternativas de sostenibilidad ecológica económica y cultural (Escobar-Mamani et al., 2020).
En lo concerniente a la información morfométrica; nos da una dimensión de descriptores más relevantes, como es el caso de longitud y grosor del peciolo, así como longitud y ancho de la hoja. De ello, se puede apreciar que C. arborea y W. mariquitae presentan hojas relativamente grandes comparadas con el resto de las especies evaluadas. Lohbeck et al. (2015), menciona que plantas con este tipo de características presentan mejor captación de luz e intercambio de gases, lo cual permite una alta capacidad fotosintética, se dice también que son indicadores de mayor vida útil de hojas y tolerante a daños por factores ambientales. Al respecto Jáuregui y Ruiz-Zapata (2011), establecen que el género Cleome dada sus características fisiológicas ha despertado interés en estudios relacionados con estructura de lámina foliar como mecanismo fotosintético.
Por otra parte Bernal et al. (2020), establece que las especies de este tipo de bioma presentan hojas pequeñas, gruesas y coriáceas, en la presente investigación este tipo de características fueron propias de la especie W. mariquitae. Esta diferencia de medidas morfométricas encontradas para las especies mencionadas nos da una idea de la complejidad y gran diversidad que este tipo de ecosistemas puede albergar, el cual nos invita a seguir estudiando para comprender mejor su dinámica de crecimiento, la población que lo conforma y el potencial uso que podrían tener bajo sistemas de producción. En este sentido Lohbeck et al. (2015), menciona que es importante identificar los múltiples rasgos de las especies ya que de esta manera se establece que estrategias toman las plantas para avanzar en temas de adaptación ante gradientes ambientales y con ello comprender mejor los procesos de regeneración y desarrollo. Información que puede ser relevante para procesos de domesticación e implementación de nuevos sistemas agroecológicos que busquen suplir las necesidades locales, tal como se muestra en la investigación realizada por Quiñones et al. (2020), quienes luego de caracterizar la vegetación nativa la seleccionaron y priorizaron de acuerdo a los rasgos funcionales para posteriormente implementar diseños agroecológicos de sistemas silvopastoriles, brindando una alternativa forrajera en épocas donde el alimento para el ganado es escaso.
Al igual como se mencionó anteriormente en el análisis de componentes principales para las cuatro especies evaluadas se pudo apreciar que los rasgos morfológicos con mayor presencia fueron ancho de la hoja, largo de la hoja y longitud del peciolo principal; es decir, variables que tienen que ver con el área foliar, factor relevante que las plantas han considerado como estrategia para mejorar la captura de energía lumínica para procesos fotosintéticos. Gómez et al. (2021), indican que la superficie o área foliar de una planta se relaciona específicamente con la cantidad de luz que una especie puede recolectar, puede ser además un indicador del estrés ambiental. Castillo et al. (2014), mencionan que esta característica se emplea como indicador de procesos fisiológicos y morfológicos que dependen de la actividad fotosintética, así como la respiración, la división celular y otros factores.
Cervantes et al. (2017) indican que, en el análisis de contribución de variables para identificar rasgos morfológicos, incluir muchas de las mismas puede afectar la clasificación, se deben considerar descriptores que mayor aporte realicen a este tipo de estudios. De este modo, en el análisis de correspondencia múltiple bajo las condiciones ambientales del sitio de estudio, podríamos mencionar que los descriptores que identifican mejor las cuatro especies fueron; base de la hoja, color de la hoja en el envés y color de la hoja en el haz. Al respecto Ureta-Leones et al. (2018), establecen que los caracteres o atributos tanto cualitativos como cuantitativos de una planta son resultados de las condiciones ambientales y características genéticas que se expresan según el caso.
Los resultados de esta investigación son importantes ya que a diferencia de muchas especies actualmente no se cuenta con estudios de caracterización morfológica de C. arborea; W. mariquitae; V. triphyllum y L. subseriata, probablemente obedezca a la bioeconomía presente en la región, la cual concentra sus esfuerzos económicos y de investigación generando paquetes tecnológicos para especies introducidas y con alto valor comercial como es el caso de pino o eucalipto de las que se obtiene materia prima para diversos productos para la industria maderera y de papel (Pérez Reyes et al., 2020; Rizza, 2020).
De esta manera la presente investigación se propone como una primera aproximación, para la caracterización de materiales promisorios en sistemas agroecológicos. Los resultados obtenidos acerca de la identificación morfológica de hojas facilitarán el seguimiento. Permitirá proponer estrategias de conservación y uso sostenible de los recursos naturales como es el caso de sistemas agroforestales y constituirá una forma de recuperar paisajes degradados por acciones agropecuarias y ambientales (Villena et al., 2019).
Conclusiones
Este trabajo contribuye a identificar los rasgos morfológicos más relevantes de cuatro especies arbustivas del bosque alto andino las cuales podrían ser potenciales en la implementación de sistemas agroecológicos que coadyuven a la sostenibilidad de zonas perturbadas por acciones agropecuarias. Y sobre todo fortalece a la interacción entre el ser humano y el medio ambiente en los Andes.
Para la especie C. arborea los descriptores cuantitativos que más aportaron fueron ancho de hoja y longitud del pecíolo. Con relación a las variables cualitativas que mayor aporte hicieron fueron ápice de la hoja, forma de la hoja, color de la hoja en el haz y color de la hoja en el envés.
Para L. subseriata se estableció que los descriptores cuantitativos más relevantes fueron largo de la hoja, ancho de la hoja y longitud del peciolo principal, mientras que las variables cualitativas más relevantes fueron base de la hoja, color de la hoja en el haz y color de la hoja en el envés.
Los descriptores cuantitativos más relevantes para W. mariquitae fueron ancho de la hoja, largo de la hoja y grosor del peciolo. Respecto a las variables cualitativas los descriptores que más aportaron fueron color de la hoja en el envés, color de la hoja en el haz y base de la hoja.
Para V. triphyllum los descriptores sobresalientes fueron ancho de hoja, largo de hoja y grosor del peciolo. Por su parte el análisis de correspondencia señalo que las variables más destacadas fueron base de la hoja, color de la hoja en el haz y color de la hoja en el envés.

