











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
El tubérculo de ñame (Dioscorea sp.) constituye un componente vital del sector agrícola en las zonas tropicales y subtropicales en el mundo. De las 600 especies de ñame cultivadas, la especie Dioscorea rotundata (ñame espino) es la de mayor importancia en términos de superficie cultivada, volumen y valor de producción (Asfaw et al., 2021). Sin embargo, este sistema de cultivo enfrenta grandes limitaciones agronómicas, que reducen significativamente su eficiencia económica. Lo anterior se puede atribuir a varios factores como falta del material de siembra de calidad, adaptabilidad a condiciones de estrés y susceptibilidad de los genotipos, a plagas y enfermedades (Okongor et al., 2021).
Los tubérculos de ñame tienen dos tipos de uso: consumo directo y material de plantación; considerándose como principal método de propagación vegetativa tubérculos enteros o porciones (Brice et al., 2019). Sin embargo, con este método, los tubérculos se multiplican lentamente con una relación de multiplicación de 1:3 en un período de 8 a 9 meses después de plantados. Esta situación fomenta la competencia en el mercado entre tubérculos comestibles y de semilla en los sistemas tradicionales de producción, en donde, el 25 y 50 % de la cosecha de ñame se convierte nuevamente en semilla (Dibi et al., 2016). El material de plantación no solo es escaso, también es costoso y puede representar hasta el 30 % del costo total de producción (Martínez-Reina et al., 2021). Todas estas circunstancias crean una situación de oferta de semilla insuficiente, de buena calidad genética y sanitaria, lo que limita la expansión del cultivo de ñame y la necesidad de buscar métodos de propagación alternativos que aporten a abastecer la demanda de material de plantación.
Los esquejes apicales de ñame para obtención de semilla son una alternativa prometedora y garantiza la disponibilidad de material de plantación durante todo el año (Claudius-Cole et al., 2020). Esta técnica ha sido probada con cierto éxito en D. alata y D. rotundata con registros de supervivencia del 30 al 40 % y tasa de multiplicación para esquejes de 20 cm de largo con 1-3 nudos de 1:30 con respecto a la tasa de multiplicación del sistema tradicional 3:1 (Agele et al., 2010). Esta tasa de multiplicación del ñame mediante esquejes ha sido aumentada a 1:300 empleando técnicas modernas en las que combinan producción de plántulas mediante sistema de biorreactor de inmersión temporal y sistemas aeropónicos (Balogun et al., 2014; Maroya et al., 2014). A partir de esquejes se obtiene una producción promedio por planta de 0.5 a 2 mini tubérculos de respectivamente (Wickham, 2019).
Sin embargo, la tasa de supervivencia, el enraizamiento y la producción de mini tubérculos siguen siendo generalmente bajos e inestables para ñame espino, la especie con mayor demanda en el mundo. Por lo siguiente, una alternativa técnicamente viable para resolver estos problemas es el uso de inoculantes a base de rizobacterias; estos microorganismos tienen la capacidad de colonizar los sistemas radiculares, producir fitohormonas vegetales, solubilizar fosfatos inorgánicos, fijar nitrógeno atmosférico, producir metabolitos secundarios e inducir tolerancia frente a condiciones de estrés biótico y abiótico (Chauhan et al., 2021). La inoculación de rizobacterias nativas en plantas in vitro ha mostrado efectos positivos en el crecimiento (Sánchez-López y Pérez-Pazos, 2018), asimismo, bajo condiciones de campo y semillas provenientes de segmentos de tubérculo, las rizobacterias lograron estimular la producción de tubérculos tipo exportación, generando beneficios económicos y ambientales (Sánchez et al., 2018; Sánchez et al., 2021). Por lo tanto, nuestra hipótesis sugiere que la inoculación de rizobacterias efectuará una respuesta favorable en los esquejes apicales de ñame.
En la región Caribe de Colombia, la producción de ñame espino se desarrolla en zonas de alta y baja montaña, principalmente en los departamentos de Bolívar, Córdoba y Sucre. El esquema de producción de material de siembra se desarrolla de forma intensiva, con prácticas agrícolas como la quema que conducen a la degradación de los suelos, erosión edáfica y remoción de material vegetal que han desmejorado las condiciones del suelo y por ende contribuido a la insostenibilidad ambiental y disminución de la biodiversidad en el ecosistema (Tamara et al., 2021). Por tanto, la inclusión de prácticas sustentables dentro del paquete agronómico del cultivo, como lo es, el uso de rizobacterias, representa una contribución al desarrollo sostenible del cultivo y la región (Escobar-Mamani et al., 2020). Por lo cual, la producción de ñame deberá realizarse bajo un enfoque transdisciplinario en el cual, se incluya aspectos socioculturales y ambientales que permitan comprender el impacto del uso de inoculantes en la mejora de las condiciones productivas de los agroecosistemas de ñame en las zonas de alta y baja montaña (Haller y Branca, 2020).
El ñame es una planta originaria de zonas boscosas, en donde la regulación de la intensidad lumínica contribuye a regular de manera eficiente los procesos de llenado de tubérculo y crecimiento de las plantas (Sugihara et al., 2020), por tanto, se deben emplear niveles adecuados de sombra que permitan optimizar la tuberización durante los procesos de multiplicación. En este sentido, en este estudio se evaluó la respuesta del ñame espino a la inoculación de la rizobacteria Bacillus licheniformis-DSC1 bajo diferentes niveles de sombreamiento y dosis de inóculo, con el fin de mejorar la sobrevivencia, crecimiento y tuberización de esquejes apicales.
Materiales y métodos
Sitio de estudio
El estudio se realizó bajo condiciones de casa de malla en el Centro de Investigación Turipaná (C.I.) de la Corporación Colombiana de investigación Agropecuaria- AGROSAVIA, ubicado en el km 13 en el municipio de Cereté (Córdoba, Colombia) a una altitud de 13 metros sobre el nivel del mar. Las coordenadas geográficas son 8°50’79” N y 75°47’58” W. La zona de estudio presenta características agroecológicas acordes a un bosque seco tropical (Bs-T), con una temperatura media anual de 27.4 °C, humedad relativa del 84 %, precipitación media anual de 1346.1 mm y brillo solar anual de 2108.2 horas (Palencia et al., 2006).
Material vegetal
El material vegetal utilizado en este estudio fue la accesión 0307-49SB de la especie D. rotundata del banco de germoplasma de ñame de AGROSAVIA, las cuales fueron propagadas mediante el protocolo (código: GA-O-13) y aclimatadas en casa de malla para la obtención de plantas madres (Figura 1a).A partir en la sexta semana de crecimiento se obtuvieron esquejes apicales con 2 a 3 nudos y una longitud de 7 a 10 cm de largo, los cuales fueron despojados de sus hojas para reducir la transpiración y una de sus hojas de la parte superior se dejó en su lugar para permitir la respiración, así como la actividad fotosintética (Figura 1b). Seguidamente, fueron sembrados en bandejas plásticas, las cuales, contenían 85 % de aluvión, 10 % de arena y 5 % de materia orgánica (Figura 1c). Este sustrato presentó las siguientes características físico-químicas: textura arenosa franco, pH 6.61, conductividad eléctrica 0.22 dS m-1, capacidad de intercambio catiónico 6.48 cmol (+) k-1, materia orgánica 0.49 g 100 g-1, fósforo disponible 4.05 mg kg-1, potasio <0.09 cmol (+) kg-1. El sustrato para la siembra de los esquejes fue desinfectado con el producto West Terrasafe® (5.0 mL L-1).

Inoculante microbiano
La rizobacteria empleada en este estudio B. licheniformis-DSC1 es nativa de la rizosfera de los cultivos de ñame en la región Caribe de Colombia y fue seleccionada de las colecciones del banco de trabajo del Laboratorio de Microbiología Agrícola del C.I Turipaná-AGROSAVIA, dada su alta capacidad para estimular el crecimiento de plantas in vitro de ñame y producir fitohormonas bajo condiciones de laboratorio en los ensayos realizados por Sánchez-López y Pérez-Pazos (2018). El inoculante de esta rizobacteria fue producido durante 15 horas en un agitador orbital (Labnet Orbit™ 1900, USA) a 150 rpm, a partir de cultivos puros y en caldos líquidos con fuentes agroindustriales optimizadas para su multiplicación (Luna-Castellanos y Sánchez-López, 2022). Siendo aplicado este inoculante a las plantas con una población bacteriana correspondiente a 108 UFC mL-1.
Establecimiento y manejo del experimento
En este estudio se evaluó el efecto de los factores: niveles de sombreamiento (0 %, 65 % y 80 %) y dosis de inoculación (2 y 6 mL). La combinación de los factores fue dispuesta en un diseño de bloques completos al azar con arreglo factorial, con nueve tratamientos y tres repeticiones cada uno. Cada tratamiento fue evaluado sobre una población de 24 esquejes apicales dispuestos en una bandeja plástica. Los tratamientos evaluados se detallan en la Tabla 1.
Tabla 1 Descripción de tratamientos.
| Tratamientos |
| T1: Sin inoculación y 0 % de sombra |
| T2: Sin inoculación y 65 % de sombra |
| T3: Sin inoculación y 80 % de sombra |
| T4: 2 mL de B. licheniformis-DSC1 y 0 % de sombra |
| T5: 2 mL de B. licheniformis-DSC1 y 65 % de sombra |
| T6: 2 mL de B. licheniformis-DSC1 y 80 % de sombra |
| T7: 6 mL de B. licheniformis-DSC1 y 0 % de sombra |
| T8: 6 mL de B. licheniformis-DSC1 y 65 % de sombra |
| T9: 6 mL de B. licheniformis-DSC1 y 80 % de sombra |
Fuente: elaboración propia.
El factor sombra fue empleado como una estrategia para facilitar la aclimatación de los esquejes de ñame. Durante el periodo experimental se utilizó polisombra al 65 %, con la cual se registró una radiación de 1100 umol ms-1, temperatura media de 29 °C y humedad relativa del 77 %. Además, con polisombra al 80 %, la temperatura media fue de 30.06 °C, humedad relativa promedio del 80.66 % y radiación de 790 umol ms-1. Sin el uso de polisombra (0 %), la entrada de luz registrada fue de 1660 umol m/s, temperatura media de 31.46 °C y humedad relativa del 78.66 %. La radiación (Photosynthetically Active Radiation, siglas en inglés-PAR) fue determinada con un Ceptómetro AccuPAR LP-80® (Decagon Devices Inc., Pullman, WA, USA).
La inoculación microbiana de los esquejes apicales se realizó dos días después de la siembra (DDS) y transcurridos 45, 95 y 130 DDS, el inoculante fue dispuesto con una jeringa en las dosis indicadas al pie de cada planta. La fertilización química se realizó de forma conjunta con estas tres últimas aplicaciones del inóculo bacteriano. Se empleó el fertilizante comercial Triple 15® en dosis total de 10 g planta-1 y su aplicación se realizó de forma fraccionada: 20 % en la primera aplicación, 30 % en la segunda dosificación y el 50 % restante en la última aplicación. La dosis del fertilizante fue calculada según los requerimientos del cultivo y análisis de las propiedades químicas del sustrato. Los esquejes fueron regados de forma manual con una frecuencia diaria. El manejo agronómico de los esquejes consistió en el control manual de malezas y monitoreo de insectos. Se realizaron aplicaciones de forma preventiva de los fungicidas Azoxistrobina+Difeconazole (2.0 mL L-1), Mancozeb+Oxicloruro de cobre (2.0 g L-1) y Metalaxil-M+Mancozeb (2.0 g L-1) con una frecuencia quincenal. Cada molécula fúngica fue aplicada de forma independiente.
Recopilación de datos
Las mediciones se realizaron a partir del día 20 después de la siembra de los esquejes. En cada unidad experimental se registró la sobrevivencia de los esquejes. A partir de los 90 a los 210 DDS, se realizaron muestreos destructivos con una frecuencia mensual. En cada fecha de muestreo (90, 120, 150, 180 y 210 DDS) fueron seleccionadas al azar tres plantas/tratamiento, se registró peso fresco de la parte aérea, radicular y número de mini tubérculos planta-1. El peso seco de los órganos vegetativos se obtuvo después de 48 horas de secado en estufa (Memmert, UN30, Alemania) a temperatura de 70 °C ± 2 °C. Los datos de peso fueron obtenidos con una balanza (OHAUS CORPORATION, V11P6, USA). Con base en los datos de peso fresco y seco, se calcularon los siguientes índices fisiológicos:
Materia seca (MS%): se determinó con base en el peso fresco y seco de la planta (Valerga et al., 2019), utilizando la fórmula de la ecuación 1:
Dónde: PFP: peso fresco de la planta; PSP: peso seco de la planta.
Contenido relativo de agua (CRA%): se calculó mediante la ecuación 2 propuesta por Gil et al. (2014).
Dónde: PF: peso fresco de la planta, PS: peso seco total de la planta.
Tasa de crecimiento absoluta (TAC): la determinación se realizó con base en la ecuación 3 (Aguilar-Carpio et al., 2018).
Dónde: W2 (peso seco final) y W1(peso seco inicial): peso seco de la planta, t2 (tiempo final) -t1 (tiempo inicial): días transcurridos entre muestreos sucesivos; sus unidades corresponden a g día-1.
Tasa de crecimiento relativo (TCR): indica la ganancia en biomasa con respecto al tiempo (Aguilar-Carpio et al., 2018). Se estimó con la ecuación 4:
Dónde: Ln W2 y Ln W1: logaritmo natural de la cantidad de biomasa seca final e inicial, de la planta completa u otros órganos individuales en un intervalo de tiempo. t2 y t1: momento final e inicial de tal periodo; siendo sus unidades gg-1·d-1.
Proporción de tallo (PDT): es la relación de biomasa seca de tallo con respecto a la biomasa total seca de la planta (Di Benedetto y Tognetti, 2016). Se estimó a partir de la ecuación 5 y se expresa en g (tallo) planta-1.
Índice de cosecha (IC): indica la cantidad de materia seca acumulada en el órgano de interés agronómico, en relación con la biomasa total de la planta (Escalante y Kohashi, 1992). Se empleó la ecuación 6 para su determinación:
Dónde: PSMT: peso seco del mini tubérculo, PSP: peso seco total de la planta.
Índice de área foliar (IAF): se evaluó con una frecuencia mensual desde los 90 hasta los 180 DDS, utilizando un Ceptómetro AccuPAR LP-80® (Decagon Devices Inc., Pullman, WA, USA).
Pigmentos fotosintéticos: la determinación se realizó desde los 90 a los 210 DDS con una frecuencia mensual, utilizando el método de DMSO (Dymethyl-sulfoxide) (Barnes et al., 1992). En cada fecha de muestreo (90, 120, 150, 180 y 210 DDS) se extrajeron discos de hojas de 0.5-0.8 cm de diámetro en cada tratamiento, los cuales fueron depositados en tubos eppendorf de 1 mL y almacenados en neveras de icopor para su refrigeración y transporte al Laboratorio de Microbiología Agrícola del C.I. Turipaná-AGROSAVIA. En el laboratorio, los discos de hojas fueron colocados en tubos de ensayo con 1 mL del reactivo DMSO, posteriormente fueron colocados durante 2 horas en una estufa (Memmert, UN30, Alemania) a 80 °C. Transcurrido este tiempo, se descartó el disco de hoja y el líquido resultante se agregó en la cubeta de medio del espectrofotómetro. Posteriormente, se midió la absorbancia a 665, 649 y 480 nm en un espectrofotómetro (Genesys™ 10S UV-Vis, Thermo Scientific®, USA), empleando como blanco el DMSO. La concentración de clorofilas (siglas en inglés Chl: chlorophyll) se calculó mediante las ecuaciones 7, 8 y 9:
Dónde: x + c = xantofilas + carotenos.
Porcentaje de tuberización: se estimó a partir del conteo del número de plantas con presencia de mini tubérculos formados al momento de la cosecha.
Análisis estadístico
Los datos obtenidos fueron analizados en el software SAS Enterprises (Cary, NC, USA). Inicialmente, fueron comprobados los supuestos del modelo, seguidamente se realizó análisis de varianza y pruebas de comparación de medias de Tukey (P<0.05) para detectar efectos significativos entre tratamientos. Además, se graficaron las dinámicas de las medias de las variables evaluadas y se ajustaron por medio de un análisis de regresión para obtener los modelos de mejor ajuste estadístico. Las gráficas fueron realizadas en el programa Microsoft Excel®.
Resultados
Sobrevivencia de esquejes
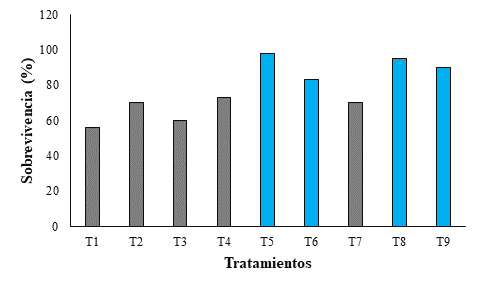
Los esquejes inoculados con B. licheniformis-DSC1 y sombra del 65 y 80 % presentaron tasas de sobrevivencia mayores al 70 % (Figura 2). La tasa de sobrevivencia más alta con relación a la sombra fue obtenida con 65 % (Figura 2). Respecto a la dosis, el mayor porcentaje de supervivencia (98 %) se presentó con 2 mL (Figura 2).
Efecto de B. licheniformis-DSC1 sobre índices de crecimiento y pigmentos fotosintéticos
La acumulación de materia seca (MS %) fue mayor con 65 % de sombra (Figura 3a). La MS no mostró variaciones significativas entre plantas inoculadas o sin inocular (P≥0.05), es decir, que el factor dosis no ejerció efecto en este parámetro. Las plantas presentaron mayor acumulación de MS a los 90 DDS con disminución progresiva hasta 180 DDS (Figura 3a). En relación con el contenido relativo de agua (Figura 3b), el factor sombra no ejerció efecto significativo (P≥0.05). La respuesta fue significativa (P≤0.05) con el uso de las dosis de inóculo, aunque entre estas no se hallaron diferencias estadísticas (P≥0.05).
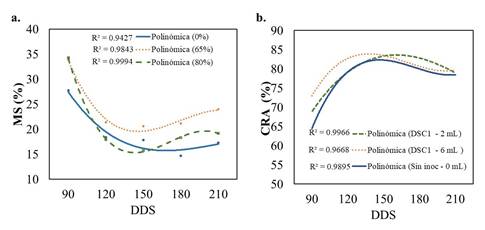
Figura 3 Respuesta de esquejes de ñame a la sombra e inoculación de B. licheniformis-DSC1. a. Comportamiento de la acumulación de Materia Seca (MS %). b. Contenido relativo de agua (CRA) en plantas inoculadas con la rizobacteria DSC1. Fuente: elaboración propia.
En cuanto a la tasa absoluta de crecimiento (TAC) y relativa (TCR) presentaron un incremento significativo del 13.8 % con sombra del 65 y 80 % en comparación con las plantas expuestas a la radiación (Figura 4a y 4b). Además, las plantas inoculadas mostraron mayor ganancia de gramos por día (entre 3 y 5 %), con respecto a las no inoculados. La dosis 6 mL tuvo mayor efecto sobre la TAC al combinarse con 80 % de sombra (Figura 4a). Con ambas dosis de inoculante se incrementó significativamente la TCR (Figura 4b). Además, la proporción de tallo (PDT) solo fue afectada por la sombra (Figura 4c), sin embargo, no se presentaron diferencias estadísticas significativas entre esquejes con y sin inóculo para este índice. La correlación (Figura 4d) entre proporción del tallo e índice de cosecha muestra una relación negativa, por tanto, PDT podría ser un factor determinante en el rendimiento de mini tubérculos.
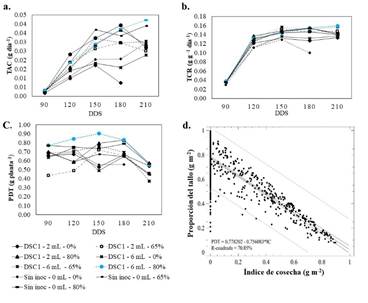
Figura 4 Efecto de niveles de sombreamiento e inóculo sobre tasas de crecimiento de plantas de ñame. a. Tasa absoluta de crecimiento (TAC). b. Tasa relativa de crecimiento (TRC). c. Proporción de tallo (PDT). d. Correlación entre proporción de tallo (PDT) e índice de cosecha (IC). Fuente: elaboración propia.
Los índices de crecimiento MS, CRA, TAC, TCR y PDT evaluados bajo los factores sombra y dosis de inóculo se ajustaron con mayores coeficientes de determinación a un modelo polinómico de tercer grado (tipo sigmoide). Este tipo de modelo de crecimiento en plantas permite determinar la asíntota superior donde se maximiza la respuesta del vegetal ante la interacción genotipo por ambiente, además indica el punto de inflexión, en donde la aceleración del proceso de crecimiento cambia de signo, pasando de una velocidad creciente a una decreciente.
En cuanto al índice de área foliar (IAF) de D. rotundata accesión 0307-49SB, los resultados demostraron que este parámetro, no estuvo influenciado por el nivel de radiación permeado por la sombra o la inoculación (P≥0.05). No obstante, a los 210 DDS las plantas bajo sombra al 65 % tuvieron mayor valor de IAF (5.58 ±2.17) que las plantas con 80 % de sombra (4.79±2.17), aunque entre estos valores no se registraron diferencias estadísticas significativas.
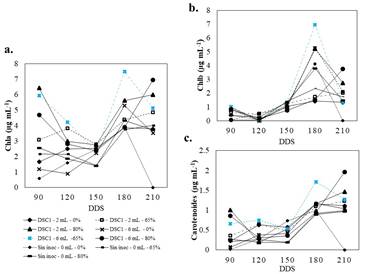
Los niveles de sombreamiento (65 y 80 %) ejercieron efecto significativo (P≤0.05) sobre la producción de pigmentos fotosintéticos (Figura 5a, 5b y 5c). La inoculación de B. licheniformis-DSC1 en dosis de 2 y 6 mL indujo una mayor producción de Chla y carotenoides (Figura 5a y 5c). Por su parte, la acumulación de Chlb (Figura 5b) no fue influenciada por la presencia de sombra o inóculo (P≥0.05). La máxima acumulación de pigmentos fotosintéticos se alcanzó a los 180 DDS con dosis de 6 mL y 65 % de sombra para Chla y carotenoides (Figura 5a y 5c). La máxima acumulación de Chlb fue obtenida con 6 mL de B. licheniformis-DSC1 y 65 % de sombra (Figura 5b).
Efecto de sombra e inoculación sobre la tuberización
La producción de semilla de ñame a partir de esquejes apicales estuvo influenciada significativamente por la combinación de sombra y dosis de inóculo. En general, las plantas bajo sombra al 65 % expresaron un IC de 0.236±0.02, duplicando en IC a las plantas bajo sombra al 80 % (0.132±0.015) y 0 % (0.095±0.017) (Figura 6a). La mayor producción de mini tubérculos representados en IC fue obtenida a los 210 DDS con sombra del 65 % (Figura 6a), entre dosis de inóculo no se hallaron diferencias destacables (Figura 6b), por lo tanto, la inoculación de las plantas con una dosis de 2 mL permite obtener un buen porcentaje de tuberización y optimizar la producción del inóculo.
Los esquejes presentaron raíces adventicias y desarrollaron raíces tuberosas. El porcentaje de tuberización se presentó en un rango entre el 20 al 100 %. La aplicación de sombra e inóculo duplicó el porcentaje de tuberización con respecto a los esquejes no inoculados o sin sombra.
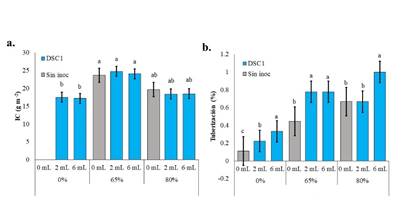
Figura 6 Crecimiento radicular y tuberización de esquejes de ñame espino. a. Índice de cosecha del cultivo (IC). b. Porcentaje de tuberización. DDS: 210. Los valores con letras diferencias indican diferencias estadísticas significativas al nivel de (P≤0.05), por el procedimiento de comparación múltiple de Tukey. Las barras indican media más el error estándar individual para cada tratamiento. Fuente: elaboración propia.
Discusión
A partir de los hallazgos encontrados en este estudio se logró evidenciar que la producción de semillas a partir de esquejes apicales constituye una estrategia viable de multiplicación para ñame espino. La inoculación de B. licheniformis-DSC1 y uso de sombra mejoro la sobrevivencia e indujo un mayor porcentaje de tuberización.
El efecto positivo de los niveles de sombreamiento (65 y 80 %) sobre el crecimiento, se encuentra estrechamente relacionado con el origen genético y domesticación del ñame espino, el cual es un híbrido homoploide procedente de la especie de sabana D. abyssinica y la especie forestal D. praehensilis (Sugihara et al., 2020) y su distribución ocurrió en el ecotono de bosque/sabana adaptándose a diferentes regímenes lumínicos (Scarcelli et al., 2019). De acuerdo con Onwueme y Johnston (2000), la aplicación de sombra sobre el cultivo de ñame genera incrementos del área foliar y cambios en el grosor de la epidermis de las hojas. El cambio en el tamaño de las hojas podría deberse a medidas compensativas como respuesta a la presencia de sombra; lo cual genera variaciones en el contenido de pigmentos fotosintéticos e incrementa la clorofila en plantas bajo sombrío, lo cual pude ser consistente con los resultados de Johnston y Onwueme (1998) en diferentes raíces y tubérculos, incluidos el ñame, donde se observó un incremento significativo de pigmentos fotosintéticos en plantas bajo sombra en comparación con plantas no sombreadas. Este incremento de clorofila podría considerarse una importante adaptación a las condiciones de sombra, relacionada con la capacidad de la planta para aprovechar limitadas cantidades de luz incidente. En este estudio, la máxima acumulación de pigmentos fotosintéticos ocurrió a los 180 DDS y su posterior descenso puede considerarse un signo de precocidad en los esquejes plantados. Las plantas de ñame con baja precocidad muestran retraso en la senescencia de las hojas, causado por estrés abiótico, el sostenimiento de hojas verdes reduce la elongación y turgencia de las hojas, retardando la translocación de carbohidratos a la raíz (Rolando et al., 2015), al presentarse amarillamiento de las hojas se incrementa la acumulación de carbohidratos en la raíz, lo que es consistente con la reducción de los pigmentos fotosintéticos a los 210 DDS y el incremento del índice de cosecha (IC) en el mismo periodo de tiempo.
En investigaciones de Primananda (2020), resalta el efecto beneficioso del uso de niveles regulados de intensidad lumínica (50-60 %) para optimizar la producción del cultivo, mejorar el comportamiento fisiológico de la planta y la estructura morfológica y anatómica de las hojas. El ñame es una planta que requiere intensidad lumínica moderada para reducir la alta fotorrespiración; producto de su metabolismo como planta C3 que incrementa su adaptación a condiciones ambientales estresantes (Cornet et al., 2007). Bajo estas condiciones de sombra las plantas de ñame desarrollan hojas más grandes para mejorar la captura de luz, estas variaciones también son visibles en el aumento de la longitud del tallo e incremento de la proporción de tallo (PDT), aunque el crecimiento radicular no responde directamente a la luz, las plantas con cambios morfológicos en el tallo y mayor PDT tienden a desarrollar raíces menos gruesas para mitigar la perdida de nutrientes por demanda de traspiración y conductancia estomática (Poorter et al., 2019). El aumento de la PDT puede incrementar el nivel de auxinas y ácido giberélico, retrasando la tuberización e induciendo el crecimiento vegetativo (Mariana y Hamdani, 2016), consistente con la relación negativa hallada entre el PDT y el IC en este estudio. Los resultados indican que el PDT disminuye a los 210 DDS y es menor en las plantas inoculadas con 2 y 6 mL de B. licheniformis-DSC1, con 0 y 65 % de sombra. La relación negativa entre el PDT y el índice de cosecha del ñame se ha documentado previamente en otras raíces y tubérculos. Investigaciones realizadas por Ramírez et al. (2017) determinaron la correlación negativa entre el área foliar y el almacenamiento de materia seca en tubérculos de camote, la cual se considera un aspecto positivo en el cultivo de tubérculos con fines productivos. La tuberización precoz se asocia con la translocación temprana de carbohidratos a la raíz, por lo que se evita la perdida de carbohidratos para mantenimiento durante la respiración en las hojas (De Souza et al., 2017).
Al parecer, la intensidad lumínica y las hormonas vegetales constituyen un factor determinante en los procesos de multiplicación de mini tubérculos a partir de esquejes. Al respecto, Agele et al. (2010), determinaron que la aplicación de ácido indolbutírico (IBA) al 1 % en esquejes del genotipo TDr 93-49 mejoró en 70 % el enraizamiento y en un 37.62 % la tuberización. En este sentido, Oyetunji y Afolayan (2014), encontraron que la utilización de casa de malla para aclimatación de esquejes ayuda a mejorar la sobrevivencia, contenido de clorofila y agua y el peso de los mini tubérculos.
El uso de rizobacterias en procesos de multiplicación de ñame a partir de esquejes, hasta la fecha, ha sido limitado dado el escaso desarrollo de investigaciones en cultivos de subsistencia como el ñame, por lo tanto, los resultados obtenidos en este estudio constituyen un primer reporte y permiten conocer los efectos de la inoculación en la fisiología de la planta. En todos los parámetros medidos se observó que la inoculación con B. licheniformis-DSC1, mejoraba la respuesta de los esquejes. Informes previos indican que inocular con cepas endófitas de Bacillus acelera la formación de tubérculos de papa en condiciones in vitro y ex vitro (Nookaraju et al., 2011). Los taxones bacterianos del cual hace parte el género Bacillus promueven la tuberización a través de la liberación de nutrientes del suelo, la producción de fitohormonas y la inducción de cambios en la fotosíntesis (Guo et al., 2020).
El mecanismo de acción de B. licheniformis sobre la inducción de la tuberización podría deberse a la segregación de la enzima ACC desaminasa (1-aminociclopropano-1-carboxílico) y la utilización de ACC como fuente única de nitrógeno mitigando el efecto adverso del etileno sobre la reducción de la longitud radicular y posterior diferenciación (Chookietwattana y Maneewan, 2012). El género Bacillus también puede secretar especies reactivas de oxígeno (ROS) que regulan la biosíntesis de ácido giberélico promoviendo la tuberización en condiciones in vitro (Kim et al., 2007). Reportes previos indican que los inóculos de B. licheniformis, pueden perdurar de cuatro a cinco semanas en el suelo después de un tratamiento único y el microorganismo puede ser detectable a los 50 días después de la inoculación, promoviendo de manera sinérgica el crecimiento vegetal (Lim et al., 2011), como se observó en los tratamientos inoculados en este estudio.
Los resultados de esta investigación sugieren que la perspectiva futura en los esquemas de multiplicación en ñame espino deberá incluir la utilización de esquejes apicales mantenidos bajo niveles regulados de sombra e inoculantes a base de B. licheniformis-DSC1, como estrategia para facilitar la disponibilidad del material vegetal en las diferentes épocas del año a los agricultores y por ende ayudar a mejorar la eficiencia económica del cultivo de ñame en la región Caribe de Colombia.
Conclusiones
La inoculación de esquejes apicales de ñame espino con B. licheniformis-DSC1 bajo condiciones de casa de malla, incrementó en un 42 % la tasa de supervivencia y el contenido de agua en la planta. Asimismo, las plantas inoculadas bajo sombrío registraron mayores tasas de tuberización en comparación con plantas no inoculados. La utilización de esquejes apicales e inoculantes microbianos a base de B. licheniformis-DSC1 constituyen una estrategia exitosa para mejorar la disponibilidad de material de siembra. Adicionalmente, esta estrategia de multiplicación contribuirá a obtener un mayor número de semillas y a mejorar la calidad en cada ciclo productivo, por ende, ayudará a mejorar los niveles de calidad de vida socioeconómicos de los agricultores y salvaguardar la seguridad alimentaria. La incorporación de bioinoculantes en el esquema de producción de ñame contribuirá a la recuperación de los suelos y conservación de la diversidad biológica y cultural en las regiones productoras, fomentando la sustentabilidad ecológica.

