











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
El Ñame (Dioscorea spp.) es importante para sostener la seguridad alimentaria en zonas tropicales y subtropicales del mundo. Los tubérculos son fuente importante de carbohidratos, proteína y fibra (Adjebeng et al., 2021; Worojie et al., 2021). De 682 especies reconocidas en el mundo, solo D. alata (ñame criollo), D. bulbifera (ñame papa), D. cayenensis (ñame amarillo), D. dumetorum (ñame trifoliado), D. esculenta (ñame ñampi) y D. rotundata (ñame espino) se cultivan como alimento (Pachakki et al.,2021; Waris et al.,2021).
Los tubérculos del ñame son de valor económico en los sistemas alimentarios y culturales de la región Caribe de Colombia, donde se producen 292.970 t (0.39% de la producción mundial, FAOSTAT, 2020). En esta región se siembra 92% del área total nacional, de la cual 45.47% corresponde a los departamentos de Bolívar y Sucre con las especies criollo y espino (AGRONET, 2018). Sin embargo, los rendimientos del cultivo en estas zonas productoras se han reducido por el aumento constante de plagas, lo que ha disminuido la calidad del tubérculo, su productividad y la eficiencia económica del cultivo.
El aumento de la presión de una plaga en sus diferentes etapas fenológicas ha inducido la aparición de enfermedades y causado deterioro de la calidad fisiológica y genética de los materiales de ñame. En la región Caribe se ha reportado la presencia del hemíptero Parafurius discifer (Stäl, 1860) en D. rotundata variedad “Botón” y “Alemán”, y D. alata var. “Oso” y “Diamante 22” (Álvarez et al., 2013). En el departamento de Sucre se ha registrado alta presencia de insectos vectores de la familia Cicadellidae como Oncometopia clarior (chicharrita o saltahojas); insecto responsable de la transmisión del Virus del mosaico suave del ñame (YMMV) que causa pérdidas considerables en la producción al reducir tamaño y calidad de tubérculos, y además causa daños indirectos al alimentarse de savia de tejidos, con un debilitamiento general (Álvarez et al., 2017). Diagnósticos fitosanitarios realizados por el Instituto Colombiano Agropecuario (ICA) en ñame han evidenciado al insecto plaga Diaspidiotus sp. Hemiptera Diaspididae, el cual, en poblaciones altas puede causar la muerte de plantas (ICA, 2018). En follaje se observa perforaciones causadas por larvas, de las cuales se desconoce su identidad taxonómica (Korada et al., 2010). En informes recientes la Corporación colombiana de Investigación Agropecuaria-Agrosavia ha recopilado datos de campo que indican la presencia de artrópodos barrenadores de tallo (Xystus sp) en ñame espino (Agrosavia, 2021), sin embargo, se desconoce qué materiales de ñame son atacados por este artrópodo, su biología y su potencial de daño en el sistema productivo.
A pesar de que en los últimos años se han realizado investigaciones preliminares en torno a insectos asociados al cultivo de ñame, existe un vacío de conocimiento con relación a cuáles son las plagas que afectan los diferentes órganos vegetativos de las plantas en la zona. Así mismo, se desconoce su distribución, hospederos, aspectos biológicos y ecológicos. Igualmente, no hay conocimiento de las etapas fisiológicas de la planta más susceptible al ataque de insectos, ni de umbrales de daño económico de estas plagas.
El sistema productivo de ñame es desarrollado bajo prácticas de manejo intensivas que generan alto deterioro a nivel ambiental y conllevan a la perdida de la biodiversidad y a la generación de desequilibrios en las redes tróficas (Tamara et al., 2021), por tanto, se hace necesario desarrollar modelos de producción de ñame con enfoque transdisciplinario y sostenible que contribuyan al mejoramiento de la calidad ambiental, social y económica de la región (Escobar-Mamani et al., 2020; Haller y Branca, 2020).
En el presente estudio se realizó un monitoreo de insectos en cultivos de ñame criollo y espino en las zonas productoras de Bolívar y Sucre, con el fin de identificar y caracterizar los daños causados por artrópodos en los órganos vegetativos y en los períodos de mayor susceptibilidad. El monitoreo de insectos plaga es una herramienta útil en la detección temprana del insecto y permite tomar medidas de control o manejo a tiempo, disminuyendo riesgos en la productividad y competitividad del cultivo.
Materiales y métodos
Sitio experimental
Las evaluaciones se realizaron entre julio y noviembre del 2020, en dos fincas productoras ubicadas en la subregión de Montes de María (Costa Caribe de Colombia). Según la clasificación de Köppen, es una subregión en la cual predomina el clima de sabana tropical con invierno seco (Aw) y temperatura promedio anual de 27ºC con períodos alternados de sequía (verano) y lluvia (invierno) durante todo el año (Kottek et al., 2006). La precipitación media anual es 1.500 mm y la humedad relativa está entre 75% y 85%, con incremento en el segundo semestre del año (García-Martínez y Mercado-Gómez, 2020). La subregión presenta suelos tipo inceptisoles con elevado contenido de arcilla y pH ligeramente ácido a neutro. En la Tabla 1, se detallan las zonas de estudio.
Tabla 1 Sitios seleccionados para el monitoreo de insectos en cultivos de ñame.
| Nombre del predio | Corregimiento/ Vereda | Municipio/ Departamento | Coordenadas | Altitud (msnm) | |
|---|---|---|---|---|---|
| N | W | ||||
| Cerro las Nubes (Loc1) | San Carlos/ Tierra Grata | El Carmen de Bolívar/ Bolívar | 9° 41' 48.2” | 75° 15’ 21.8” | 487 |
| Guaimaro (Loc2) | La Ceiba | Chalán/ Sucre | 9° 35' 46” | 75° 17’ 21” | 450 |
* Loc: localidad. Fuente: elaboración propia.
Material vegetal y manejo agronómico
Para realizar el monitoreo de los insectos se seleccionaron cultivos de las especies D. alata var. Diamante y D. rotundata var. Criollo con un área sembrada de 0.1 a 0.25 ha. Las variedades de ñame en ambas localidades fueron establecidas bajo monocultivo. En la Loc1, los agricultores incluyeron entre las labores de manejo del cultivo: selección y desinfección química de semillas empleando una mezcla de fungicida Ridomil Gold 68 WG® e insecticida Cypermetrina 20 EC®, controles manuales y químicos de malezas con herbicidas pre-emergentes, aplicación de fertilizantes inorgánicos en base a Urea®. Además, realizaron cuatro aplicaciones calendario del insecticida Cypermetrina 20 EC® a los 50, 90, 120 y 150 días después de la siembra (DDS). Para el establecimiento del ñame Diamante emplearon una distancia entre plantas de 0.40 m y entre surcos de 1.20 m respectivamente. Por su parte, el ñame espino criollo fue establecido con una distancia entre plantas de 0.50 m y 1.40 m entre surcos. Las variedades de ñame fueron sembradas en fecha 13/05/2020.
Las condiciones de manejo del cultivo en Loc2 fueron de tipo tradicional y sin la aplicación de agroquímicos. El ñame Diamante fue sembrado a una distancia entre plantas de 0.50 m y entre surcos de 1.20 m respectivamente. Por su parte, el ñame espino criollo fue establecido con una distancia entre plantas de 0.80 m y entre surcos de 1.20 m. Las variedades de ñame fueron sembradas el 10/05/2020. En ambas localidades se emplearon soportes de madera para facilitar el amarre y desarrollo vegetativo del ñame espino criollo. En contraste, el ñame Diamante fue sembrado en ausencia de soportes.
Monitoreo de insectos
Las evaluaciones de insectos en ñame espino criollo se realizaron en un recorrido en zig-zag seleccionando al azar diez plantas. En cada planta se inspeccionó la ausencia (0) y/o presencia (1) de adultos o daños de artrópodos en hojas o tallos del tercio inferior, medio y superior de cada planta. Para realizar el monitoreo en el área sembrada con ñame Diamante fueron seleccionados cinco puntos aleatorios de muestreo en un recorrido en zig-zag. En cada punto se hizo el registro de los artrópodos por triplicado usando un marco de 0.0625 m2 (25 x 25 cm). Los muestreos se realizaron con frecuencia quincenal en las dos variedades de ñame en ambas localidades. En la Loc1 las evaluaciones fueron realizadas a partir del 13/07/2020, mientras que en la Loc2 a partir del 10/07/2020. En la Tabla 2, se especifican las fechas de muestreo para cada localidad.
Tabla 2 Datos de muestreos realizados en cultivos de ñame.
| Mes/Fecha | DDS | |||
|---|---|---|---|---|
| Espino | Diamante | |||
| Loc1 | Loc2 | Loc1 | Loc2 | |
| Julio | 90 | 85 | 60 | 45 |
| Ago-15 | 116 | N.R | 81 | N.R |
| Ago-30 | 126 | N.R | 96 | N.R |
| Sep-15 | 140 | 144 | 110 | 105 |
| Sep-30 | 153 | 163 | 123 | 123 |
| Oct-15 | 174 | 171 | 144 | 131 |
| Oct-30 | 195 | 184 | 165 | 144 |
| Noviembre | 211 | 198 | 181 | 158 |
*N.R: no realizada por Covid-19. Fuente: elaboración propia.
Para realizar observaciones en las hojas y tallos de huevos, ninfas y otros estados inmaduros de los artrópodos se utilizó lupa de 30X. Para la colecta de adultos en cada tercio de la planta se hizo un pequeño golpe en la parte más densa del follaje con un filtro de tela para café marca Ilko® de 30 x 14 x 0.5 cm; este instrumento se utilizó para capturar los adultos, los cuales posteriormente fueron depositados en bolsas ziploc y almacenados en neveras de icopor bajo refrigeración para su procesamiento en laboratorio.
Los especímenes colectados en cada una de las localidades y variedades de ñame fueron separados teniendo en cuenta caracteres morfológicos según orden, familia y morfoespecie y conservados en alcohol al 70 % en el Laboratorio de Entomología Agrícolas del centro de investigación Turipaná de Agrosavia (N: 08°50'59”, W: 75°48'54”), empleando un estereoscopio Leica EZ4®. Posteriormente las muestras fueron enviadas para identificación taxonómica al Laboratorio de Diagnóstico Fitosanitario del ICA, Cereté, Córdoba, Colombia.
Variables respuesta
Daños Causados por artrópodos: sobre la base de reportes sanitarios previos para el cultivo de ñame en la región Caribe y revisión de literatura (Korada et al., 2010; Álvarez et al., 2013; Álvarez et al., 2017) durante la etapa de muestreo, los daños observados fueron atribuidos a algún artrópodo plaga, de tal manera que se clasificaron como: comedores de follaje (hormigas, adultos de la familia Chrysomelidae), chupadores de savia (ninfas y adultos de la familia Cicadellidae, Miridae (P. discifer) y otros especímenes del orden Hemíptera) y barrenadores de tallos y guías de la planta de ñame (larvas y adultos de Xystus sp).
Incidencia (%): Con los datos obtenidos de ausencia (0) y presencia (1) se realizó la estimación de incidencia de artrópodos en estado adulto (presencia de Cicadellidae, Chrysomelidae y P. discifer) registrados al momento de la evaluación causando daño en algún órgano vegetativo de la planta, e incidencia de daños de artrópodos (daños observados como perforaciones en hojas, tallos y raspaduras en hojas causados por artrópodos, que fueron encontradas al momento del muestreo). Se utilizó la ecuación 1 para realizar el cálculo:
Probabilidad de daño: Se determinó la probabilidad de encontrar un daño ocasionado por los artrópodos durante las fechas de muestreo, para lo cual se transformó la variable presencia (1) del daño y después se realizó una regresión logística con la ecuación 2:
Dónde: pi es la probabilidad de encontrar un daño de cualquiera de los artrópodos evaluados en un muestreo i. β0: intercepto. β1: Coeficiente de la variable variedad. β2: Coeficiente de la variable días después de la siembra (DDS). β3: Coeficiente de la variable localidad. La significancia de cada variable se comparó por medio de la prueba de Wald.
Análisis estadístico
Se realizó un análisis mediante la técnica de Análisis factorial de datos mixtos (AFDM), que incluye base de datos compuesta por variables cualitativas y cuantitativas (Pagès, 2004). Las variables cualitativas correspondieron a los factores localidades, variedades de ñame y DDS. Por su parte, las variables cuantitativas correspondieron a la ausencia (0) y presencia (1) de adultos o daños de artrópodos. El AFDM permitió la exploración simultánea de dichas variables mediante la combinación de análisis de componentes principales (ACP) y análisis de correspondencias múltiples (ACM) (Husson et al., 2010). Tras este análisis, se llevaron a cabo pruebas de comparación de medias según Tukey (P<0.05) y correlación de Pearson. Los análisis estadísticos se hicieron con el software R versión 3.4.2 (R Core Team, 2017), utilizando los paquetes FactoMineR en el cálculo de los resultados y Factoextra para la visualización de los gráficos.
Resultados
Caracterización de daños causados por artrópodos en especies de ñame
Bajo las condiciones ambientales y de manejo agronómico, en la subregión de Montes de María, durante la etapa vegetativa, los tallos y hojas de las especies de ñame pueden verse afectadas por artrópodos plagas que causan daños directos en algunas estructuras o transmiten enfermedades cuando se alimenta. Durante el período de muestreo se registraron daños causados por artrópodos del orden Coleóptera (familias Chrysomelidae y Curculionidae) y Hemíptera (Cicadellidae y Miridae). A continuación, se describen daños causados a hojas y tallos que permiten su reconocimiento:
Daños causados por artrópodos en hojas de la especie ñame
En este ítem fueron clasificados en tres subgrupos los artrópodos descrititos a continuación y presentados en la Figura 1:
Grupo 1: Denominado de escarabajos de las hojas; se encontraron cinco artrópodos causantes de perforaciones en la lámina foliar (Figuras 1a y 1b), Omophoita sp (Figura 1c), Agroiconota propinqua (Figura 1d) y tres morfoespecies de la familia Chrysomelidae pertenecientes a la subfamilia: Galerucinae, Tribu: Alticini (Figura 1e, 1f y 1g).
Grupo 2: Chicharitas o salta hojas, se los observó en el tercio superior de plantas hojas con síntomas de moteado característico de plantas afectadas por virus (Figura 1h y 1i). En estas mismas plantas fueron colectados adultos de Empoasca spp y Oncometopia sp1 (Figura 1j), los cuales han sido reportados como trasmisores del Virus del Mosaico Suave del Ñame (YMMV) del género Potyvirus.
Grupo 3: Chiches de la Familia Miridae; fueron observadas en el haz foliar (Figura 1k) puntuaciones blanquecinas a lo largo de las nervaduras de las hojas y en el envés pequeños puntos negros redondos (Figura 1l) caracteristicos de las deposiciones de P. discifer (Figura 1m).

Figura 1 Perforaciones en hojas de ñame causadas por escarabajos. a. Perforaciones en lámina foliar de D.rotundata. b. Perforaciones en hojas de D.alata. c. Omophoita sp. d. Agroiconota propinqua . e. Chrysomelidae Subfamilia Galerucinae sp1. f. Chrysomelidae Subfamilia Galerucinae sp2. g. Chrysomelidae Subfamilia Galerucinae sp3. h. Moteado en D.alata. i. Moteado en hojas de D.rotundata. j. Adultos de Oncometopia sp1. k. Punteado blanquecino en el haz de hojas de D.rotundata. l. Puntos negros (deposiciones del artrópodo) en el envés de D.rotundata. m. Adulto de P. discifer.
Daños causados por artrópodos en tallos de la especie ñame
Para este órgano, solo se observó el barrenador del tallo; las plantas afectadas presentaron inicialmente síntomas de clorosis en las hojas del tercio inferior (Figura 2a), seguidamente secamiento progresivo de los tallos afectados (Figura 2b y 2c). Al interior de las perforaciones fueron encontradas larvas y adultos de Xystus sp. (Figura 2d).

Incidencia de adultos y daños de artrópodos
El AFDM evidenció ocho variables (una cualitativa y siete cuantitativas) como factores discriminantes para comprender el comportamiento de los artrópodos evaluados en las especies de ñame.
Los resultados del AFDM indican que 30.10% de la variabilidad total de los datos puede ser explicada en tres dimensiones. La primera dimensión explica 13% de la variabilidad acumulada, la variable cualitativa DDS es la de mayor contribución a la varianza, seguida de las variables cuantitativas daños de Xystus sp y Chysomelidae (Figura 3a). La segunda dimensión explica 10.7%, donde DDS es la variable que explica en mayor proporción el comportamiento de los artrópodos evaluados; entre las variables cuantitativas se encontró que daños y presencia del hemíptero P. discifer, al igual que daños de Cicadellidae presentan una mayor correlación con esta dimensión (Figura 3a). La tercera dimensión explica el comportamiento de los datos en 6.4%; la presencia de adultos de Cicadellidae presenta mayor correlación (R2= 0.53) en esta dimensión. Se resalta que la presencia de adultos de Chysomelidae presenta correlación negativa (R2= - 0.77) en esta dimensión (Figura 3b).
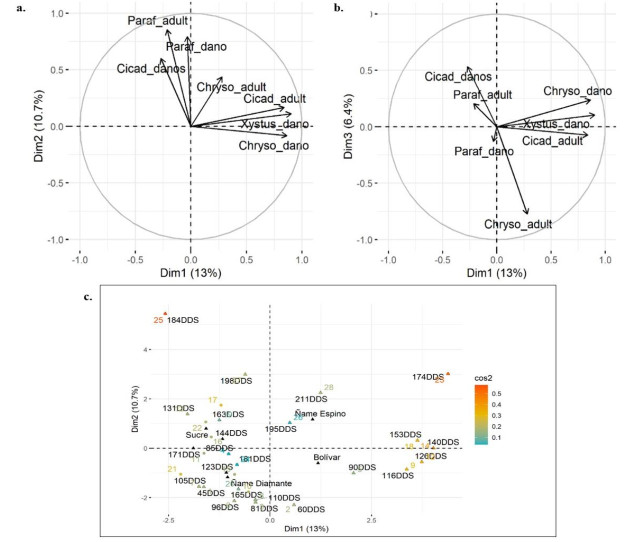
Figura 3 Circulo de correlaciones para las variables cuantitativas. a. Correlaciones variables cuantitativas dimensiones 1 y 2. b. Correlaciones variables cuantitativas dimensiones 1 y 3. c. Prueba de coseno cuadrado para variables cualitativas.
En la Figura 3c, se pueden apreciar las correlaciones de las variables cualitativas DDS, especies de ñame y localidades (Bolívar, Sucre) con las dimensiones 1 y 2. A partir de estos resultados se evinció que DDS es la variable cualitiva de mayor correlación estadistica. En relación a la dimensión 1 (Figura 3c), se puede ver que a los 174 DDS se registró la mayor correlación, esto pudo deberse posiblemente a que en ñame espino en Loc1 los artrópodos P. discifer y Chysomelidae registraron el máximo nivel de incidencia (100%) en cuanto a la presencia de daño, a partir de esta fecha de muestreo (Figura 4a). En la Figura 4a, se puede observar que la incidencia de la presencia de daño ocasionada por Xystus sp está restringida únicamente al ñame espino, a los 90 DDS se puede observar la mayor incidencia (73.33%). Por su parte, los daños de alimentación de Chysomelidae se presentan en ambas especies de ñame y son notorios desde los 90 DDS. Así mismo, es relevante mencionar que los daños de P. discifer inician en ñame espino a los 140 DDS y en Diamante a los 110 DDS respectivamente (Figura 4a). Los datos podrían indicar que P. discifer puede presentar una mayor preferencia por los tejidos foliares de las especies de D. rotundata en comparación con los D. alata.
Con respecto a la dimensión 2 (Figura 3c), la correlación de mayor significancia se presentó a los 184 DDS para ñame espino en la localidad de Sucre, lo cual puede obedecer a que los daños de P. discifer alcanzan el máximo nivel de incidencia (100%) para esta fecha de muestreo (Figura 4b). En la Figura 4b se puede observar que en la especie D. alata la mayor incidencia ocasionada por este insecto se presentó a los 131 DDS, es decir, que en esta localidad la presencia del daño alcanzó su máximo nivel 50 días antes que en la Loc1. La incidencia de los daños del coleóptero Xystus sp solo pudieron apreciarse a los 144 DDS en la especie D. rotundata. Con relación al comportamiento de la incidencia del daño de Chysomelidae en las especies de ñame, se puede apreciar en la Figura 4b que la incidencia se presentó en un nivel por debajo del 40% durante las evaluaciones realizadas, valor inferior con respecto a la Loc1.
Por otra parte, en la Figura 4b se puede advertir que bajo las condiciones ambientales y manejo del cultivo la incidencia del daño de P. discifer se registró en ñame espino a partir de los 85 DDS, lo cual podría ocasionar disminuciones considerables en el crecimiento de la planta debido a los daños foliares ocasionados. En contraste, en Loc1, este artrópodo es notorio a los 140 DDS, es decir, que bajo esta condición la aparición del insecto en el cultivo podría retrasarse 50 días en comparación con Sucre, lo cual es ventajoso para que las plantas de ñame espino puedan desarrollar el tejido foliar de forma adecuada y realizar un óptimo llenado del tubérculo.
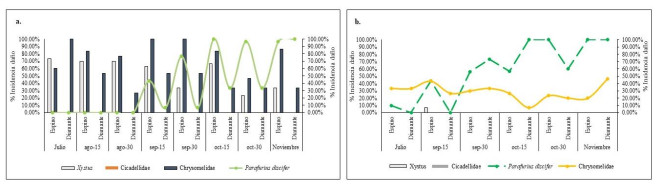
Relación entre daño de artrópodos, DDS, especies de ñame y tercios de la planta
Los resultados de la prueba de correlación de Chi Cuadrado de Pearson detallados en la Tabla 3, evidencian que hubo diferencias altamente significativas (p=<0.0001) en la incidencia del daño causada por los artrópodos respecto a los DDS en las especies de ñame. Estos resultados ratifican que la variable DDS es un parámetro determinante en el estudio de artrópodos en el cultivo de ñame, confirmando los resultados del análisis AFDM. A partir de estos resultados se podría afirmar que la etapa fenológica en ñame constituye un factor clave para comprender el comportamiento de los artrópodos y define el momento oportuno en que se debe iniciar el monitoreo de artrópodos y la aplicación de estrategias de manejo para disminuir su daño.
Tabla 3 Relación entre DDS, especies de ñame y tercios de la planta respecto al daño de artrópodos.
| Factor | Variable | Valor X2 | p |
|---|---|---|---|
| DDS | Daño Cicadellidae | 53.51 | 0.0005 |
| Daño Chrysomelidae | 96.53 | <0.0001 | |
| Daño P. discifer | 343.46 | <0.0001 | |
| Daño Xystus sp | 310.97 | <0.0001 | |
| Localidad | 553.19 | <0.0001 | |
| Especies de Ñame | Daño Cicadellidae | 5.94 | 0.0148 |
| Daño Chrysomelidae | 3.22 | 0.0727 | |
| Daño P. discifer | 7.24 | 0.0071 | |
| Daño Xystus sp | 83.73 | <0.0001 | |
| Localidad | 0.00 | >0.9999 | |
| Tercios de la planta | Daño Cicadellidae | 1.56 | 0.4576 |
| Daño Chrysomelidae | 0.89 | 0.6398 | |
| Daño P. discifer | 1.19 | 0.5515 | |
| Daño Xystus sp | 9.86 | 0.0072 | |
| Localidad | 0.00 | >0.9999 |
*Estadístico Chi Cuadrado Pearson (X2).
Fuente: elaboración propia.
Con respecto a la correlación entre especies de ñame e incidencia de daño de artrópodos, los resultados de la Tabla 3, demuestran que la incidencia del daño de artrópodos Cicadellidae. P. discifer y Xystus sp puede estar asociada de forma altamente significativa con la especie de ñame (P=≤0.01). La mayor incidencia del daño se encontró en D. rotundata cv. Espino con una frecuencia relativa (Fr) de 0.94 para Cicadellidae. 0.75 para P. discifer. y 1.00 para Xystus sp respectivamente. Por otra parte, no se detectó diferencia estadística (P≥0.05) respecto a la incidencia del daño causados por artrópodos de la familia Chrysomelidae entre las localidades y especies de ñame.
Los resultados en la Tabla 3, permiten afirmar que los artrópodos pertenecientes a las familias Cicadellidae y Chrysomelidae y la especie P. discifer pueden causar daño en el tejido foliar de las plantas de ñame indistintamente del tercio en el cual se encuentren las hojas o tallos (P≥0.05). Otro resultado, está relacionado con la selectividad de Xystus sp hacia los tercios de la planta (P=0.0072), al parecer prefiere consumir los tallos ubicados en el tercio inferior (Fr= 0.43) y medio (Fr=0.31) de D. rotundata cv. Espino (Tabla 3).
Probabilidad de daño de artrópodos Chrysomelidae, P. discifer y Xystus sp en Cultivos de ñame
Los análisis de regresión logística no detectaron diferencias estadísticas significativas entre DDS y localidades (P=0.2821) en torno a la probabilidad de daño de Chrysomelidos en ñame. Se logró estimar con el modelo que se puede presentar una probabilidad promedio de Chrysomelidos de 28.62%±0.09 para Loc1 y 19.22%±0.30 para Loc2. Asimismo, no se detectaron diferencias estadísticas entre las especies de ñame (P=0.2242). La probabilidad de presencia de daños de Chrysomelidos en ñame en la región de Montes de María registró dos períodos marcados para Loc1 y Loc2. En la Figura 5a, se puede apreciar que, en la Loc1, el primer período se presentó desde los 31 a 60 DDS y el segundo desde los 256 a 270 DDS, en ambos períodos el insecto alcanzó valores del 43.33%. Entre tanto, en la Loc2, el primer período se presentó en etapas más avanzadas de desarrollo del cultivo desde los 151 a 181 DDS y el segundo de los 211 a 240 DDS, siendo el segundo el que presentó mayor probabilidad, con valores del 90 % (Figura 5b).
En cuanto a la probabilidad de daño de P. discifer se detectaron diferencias estadísticas altamente significativas para DDS (P≤0.01). Entre las especies de ñame no se registraron diferencias estadísticas (P=0.1434). La probabilidad promedio de daño de P. discifer durante el ciclo del cultivo en Loc1 fue del 61.13%±0.37 y del 46%±0.49 para Loc2. En la Figura 8c, se puede evidenciar que en la Loc1 el artrópodo alcanzó valores del 100% desde 121 a 180 DDS y desde 240 a 270 DDS. Por su parte, en la Loc2, desde 151 a 240 DDS se registró una probabilidad por encima del 96% (Figura 5d). Teniendo en cuenta la probabilidad de daño (Figura 5c y Figura 5d), en la región de Montes de María P. discifer puede estar asociado al sistema productivo de ñame desde etapas muy tempranas, a partir de los 30 DDS. Es posible afirmar, además, que desde los 90 DDS en adelante la probabilidad de daño aumenta, lo cual podría estar relacionado con la formación continua de hojas en la planta en esta etapa fenológica del cultivo, por lo cual el insecto tendría alimento para consumir y poder incrementar su población.
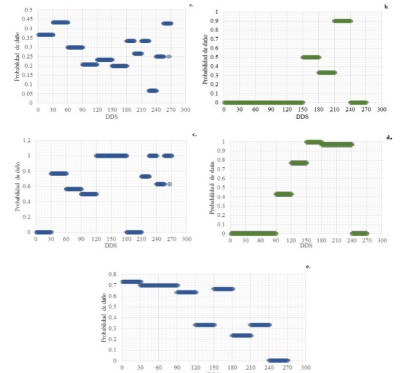
Figura 5 Probabilidad de daño de artrópodos en ñame. a. Chrysomelidos en Loc1. b. Chrysomelidos en Loc2. c. P. discifer en Loc1. d. P. discifer en Loc2. e. Xystus sp en Loc1.
La probabilidad de daño de Xystus sp. fue estimada únicamente para Loc1, debido a que en la Loc2 solo se registró por única vez este insecto a los 144 DDS (Figura 4b). Este insecto presentó una preferencia por la especie D. rotundata cv. Espino, con una probabilidad promedio de daño durante el cultivo de 48.15%±0.24 (Figura 5e). Este artrópodo puede estar asociado a la planta de ñame espino causando daños en el tallo desde las etapas iniciales de crecimiento de la planta, lo cual podría estar relacionado con el bajo nivel de lignificación de los tejidos de este órgano durante esta etapa fisiológica. En la Figura 5e se puede notar que la probabilidad del daño puede ser alta (>65%) desde la emergencia del tallo hasta los 117 DDS y se puede presentar un segundo período marcado de probabilidad de daño desde los 151 a los 180 DDS alcanzando niveles del 66.66%. El modelo permitió estimar la probabilidad de encontrar un daño causado por Xystus sp. en tallos de ñame espino dejando en evidencia la estrecha relación de Xystus sp. con las etapas fisiológicas tempranas del ñame espino.
Discusión
Los resultados obtenidos en esta investigación permitieron mostrar que durante la etapa vegetativa y las condiciones de manejo agronómico predominantes en la subregión de Montes de María, las especies de ñame D. alata y D. rotundanta son afectadas por nueve artrópodos plagas, los cuales al consumir los órganos vegetales de la planta indirectamente merman la producción eficiente de foto-asimilados requeridos para el llenado óptimo del tubérculo (Lake et al., 2015). La identificación y caracterización de daños causados por artrópodos en cultivos de ñame es escasa a nivel local e internacional, por tanto los resultados obtenidos constituyen un primer avance hacia la determinación de fluctuación poblacional, niveles y umbrales de daño y prácticas de manejo integrado para estos artrópodos.
Los daños encontrados en hojas de D. alata y D. rotundanta en forma de perforaciones circulares o alargadas estuvieron asociados a escarabajos de la familia Chrysomelidae. Esta es una familia de artrópodos herbívoros diversa y abundante del orden Coleóptera con cerca de 40.000 especies, casi todas fitófagas. Presentan amplia variedad de plantas huésped que va de briófitas hasta gimno- y angio-espermas (Iannella et al., 2021). Algunas especies se alimentan de raíces, otras de material vegetal sobre el suelo y también se alimentan de flores, polen, hojas y brotes nuevos de las plantas. Muchos crisomélidos se alimentan de forma mono y oligofágica de taxones de plantas específicos, otros, especialmente entre Eumolpinae, Cryptocephalinae y Clytrinae, pueden usar una amplia gama de plantas (Lucio-García et al., 2022).
Los escarabajos de la familia Chrysomelidae constituyen una familia de artrópodos ecológicamente importantes debido a su alta especificidad con las plantas hospederas, su estrecha asociación con la vegetación y por su gran sensibilidad a la variación microclimática (factores que se modifican gradualmente durante los períodos de lluvia y sequía). La selección del follaje diferencial intra-planta por crisomélidos se ha estudiado en varias investigaciones, sin embargo, no se ha detectado un patrón general de preferencia por hojas jóvenes u hojas maduras (Lucio-García et al., 2022). Estos artrópodos consumen el follaje y lo pueden emplear para diferentes propósitos, lo cual constituye una posible estrategia para proporcionar nichos dentro de la planta para sobrevivir en diferentes etapas de desarrollo (Fernandez & Hilker, 2007). Asimismo, la estructura del paisaje en el que crece una planta huésped puede influir significativamente en su detectabilidad y, por lo tanto, en el grado de ataque por parte del crisomélido (Ahuchaogu & Ojiako, 2021).
Durante la etapa de muestreo fueron registrados tres artrópodos de la subfamilia Galerucinae (Coleóptera: Chrysomelidae); este grupo de escarabajos saltadores y no saltadores está compuesto por unos 1.100 géneros y más de 15.000 especies (Lucio-García et al., 2019). Investigaciones realizadas en fragmentos de bosque tropical en México han revelado que los escarabajos de esta subfamilia presentan una gran dominancia frente a otros de la familia Chrysomelidae, lo cual puede deberse a que el gran número de especies que lo componen pueden sobrevivir y adaptarse a diferentes épocas climáticas del año y disponibilidad de alimento (Lucio-García et al., 2022). En cultivos de ñame en África, los daños en hojas han estado asociados al crisomélido Crioceris lívida (Asante et al., 2007), sin embargo, se desconoce la distribución geográfica, biología y dinámica de las poblaciones de este insecto y la susceptibilidad de las especies y cultivares de ñame. Reportes de investigaciones realizadas por Purushan et al. (2017) revelaron que en los cultivos de tubérculos tropicales (batata, yuca y ñame) la diversidad relativa de la familia Chrysomelidae fue superior al resto de las familias evaluadas, ratificando que estos artrópodos presentan una gran diversidad que puede ser albergada en sistemas productivos de ñame. Otra de las subfamilias de crisomélidos reportados en nuestra investigación corresponde a Cassidinae. En Colombia se han reportado 221 especies, de las cuales Agroiconota propinqua registrada durante la etapa de muestreo en esta investigación ha sido reportada como causa de daños foliares en diferentes cultivos en siete departamentos del país. En la región Caribe de Colombia fue registrada causando defoliación en cultivos de batata (Borowiec & Świętojańsk, 2015; Agrosavia, 2021).
Otro grupo importante de artrópodos registrados durante el muestreo fueron los hemípteros de la familia Cicadellidae. Cabe resaltar que, aunque fueron registrados por única vez representan una alerta temprana para prevenir posibles infecciones virales en las plantas y transmisiones masivas por la alimentación del artrópodo dentro del campo (Luo et al., 2022). Las chicharritas o salta hojas como se les conoce tradicionalmente, además de ser vectores de agentes patógenos, causan clorosis y debilitamiento generalizado en la planta al succionar la savia de las hojas para alimentarse. En la región Caribe de Colombia, trabajos realizados en ñame espino por Álvarez et al. (2017) indican que estos insectos utilizan de forma natural las hojas de la planta para alimentarse y realizar la oviposición, asimismo, manifiestan que D. rotundata presenta una alta variabilidad genética, por lo cual presenta mayor resistencia que D. alata frente a infecciones virales.
En el follaje de las especies de ñame fueron observados daños ocasionados por el hemíptero de la familia Miridae, P. discifer. Este artrópodo presenta una amplia distribución geográfica e importante función en el ecosistema ya que actúa como fitófago y depredador. Ha sido reportado en Colombia en cultivos de ñame espino y criollo (Oso y Diamante 22) (Álvarez et al., 2013). Síntomas similares a los registrados en esta investigación fueron observados en planta de cartucho y café (Carvalho et al., 2011, Arellano y Vergara, 2016). Los reportes previos encontrados hasta la fecha no han evaluado la incidencia ni la probabilidad del daño durante el ciclo de crecimiento del cultivo, por tanto, se desconocen los efectos de sus daños en el rendimiento y productividad de las plantas de ñame y aspectos importantes en torno a su biología.
En cuanto a los barrenadores del tallo, los daños estuvieron asociados a Xystus sp (Curculionidae, subfamilia Barinidae). Informes realizados por Santos et al. (2007) han evidenciado la presencia de la especie Xystus arnoldi en D. cayennensis, D. rotundata, D. alata, D. trifida y D. esculenta en el estado de Paraíba (Brasil). Los daños de este herbívoro estuvieron relacionados con el secamiento progresivo del tallo principal, comprometiendo el crecimiento y sobrevivencia de la planta. Estudios realizados en el estado de Jalisco y Michoacán en México en especies silvestres de ñame han informado de la presencia de dos nuevas especies del género Xystus correspondientes a X. gracilirostris (Champion) y X. pallidipennis (Champion). La especie X. gracilirostris ocasionó daños en semillas aéreas de ñame, mientras que X. pallidipennis en tejidos apicales de las plantas (Jones et al., 2011). En Colombia, Xystus sp fue reportado por primera vez en D. rotundata en el departamento de Bolívar en 2021 (Agrosavia, 2021). Teniendo en cuenta lo anterior se podría afirmar que las especies del género Xystus tienen una especificidad por un órgano vegetativo de ñame; asimismo, su biología y comportamiento están ligados a condiciones ambientales y tipo de hospederos. Los resultados de nuestra investigación indicaron que Xystus sp mostró una selectividad por los tallos ubicados en el tercio inferior de la planta; este hecho podría deberse a que cuando el artrópodo inicia su proceso de infestación en el cultivo de ñame espino, este período corresponde a etapas fenológicas tempranas de la planta en la cual los tallos del tercio medio e inferior están poco lignificados; además, podría deberse a que Xystus sp., tendría una baja altura de vuelo que le dificultaría causar daños en el tercio superior. Algunos estudios realizados en manglares determinaron que trampas colocadas a 1.5 m de altura capturaron mayor abundancia de Curculionidae que las colocadas a 6 m o 12 m (Gerónimo-Torres et al.,2021).
Los estudios en torno a la biología, umbrales de daño, dinámica poblacional, etapas fenológicas de mayor susceptibilidad del ñame, así como los efectos en el rendimiento por los ataques de comedores de follaje (crisomélidos, cicadélidos y P. discifer) y Xystus sp., no han sido informados a la fecha, por tanto constituyen una premisa de investigación de los países productores de ñame en Latinoamérica para salvaguardar la producción eficiente y sostenible de tubérculos de ñame.
Conclusión
A partir de evaluaciones realizadas en la etapa vegetativa de D. alata y D. rotundata en la subregión de Montes de María se identificó y caracterizo los daños causados por nueve artrópodos plagas en hojas y tallos de la planta. Los daños encontrados estuvieron relacionados con herbívoros fitófagos del orden Coleóptera (familias Chrysomelidae y Curculionidae) y Hemíptera (Cicadellidae y Miridae). Los resultados obtenidos de incidencia y probabilidad de daño constituyen una primera aproximación para la construcción de un modelo que nos permita estimar con precisión las etapas iniciales de ataque de los insectos y la severidad de los daños, con lo cual se podrán establecer planes de manejo integrado que permitan salvaguardar la producción eficiente y sostenible de tubérculos de ñame en la región Caribe de Colombia y contribuir así al sostenimiento de la economía rural de muchas familias cuya principal fuente de ingreso depende exclusivamente del ñame. Así mismo, se buscarán alternativas de manejo futuras tendientes a la conservación de la diversidad biológica de las zonas de alta montaña indispensables para proteger la sustentabilidad ecológica de la región.

