











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
La Reserva Nacional Allpahuayo-Mishana, situada en la ecorregión de selva baja, posee una gran importancia para la conservación de la biodiversidad peruana, dado el alto número de especies que se ha registrado allí. La diversi- dad vegetal de la RNAM es extraordinaria, pues reúne a más del 10% de las especies de plantas reportadas en el Perú (Brako & Zarucchi, 1993) y presenta unos de los valores más altos del mundo en diversidad de árboles, palmeras y lianas por hectárea (Vásquez & Phillips, 2000). La riqueza de vertebrados de esta reserva es también nota- ble: el número de especies de aves registradas en la RNAM, 496, supera al de áreas protegidas mucho más extensas (Alvarez et al., 2012) y equivale al 26% de las 1882 especies registradas en el país (Plenge, 2022), mientras los reptiles, con 56 especies (Pérez et al., 2017a), y los anfibios, con 68 (Pérez et al., 2017b), representan el 12 y el 11% de la riqueza peruana, respectivamente (MINAM, 2018). Con respecto a los mamíferos, la RNAM es el segundo sitio del Perú más rico en especies de murciélagos, con 65 registradas (Hice et al., 2004), y tiene una de las comunidades más diversas del Neotrópico en cuanto a pequeños roedores y marsupiales, con 37 especies (Hice & Velazco, 2012).
A diferencia de otros grupos de vertebrados, para los mamíferos terrestres medianos o gran- des, definidos como aquéllos cuyos adultos pesan 1 kg o más (e.g. Botelho et al., 2012, dos Santos & Rodrigues 2013), no se cuenta en la RNAM con evaluaciones recientes basadas en registros directos. La mayor parte de los registros de especies publicados para este grupo se basan en pieles o cráneos obtenidos de cazadores locales, huellas y entrevistas (Oversluijs, 2003; Hice & Velazco, 2012). Sin embargo, los registros directos son importantes para el manejo de un área prote- gida, pues ofrecen seguridad sobre la presencia de especies raras o poco observadas por la pobla- ción local, verifican la presencia actual y el lugar exacto de registro y confirman registros visuales y entrevistas que podrían presentar dudas.
Entre los métodos de registro directo de mamíferos terrestres, el uso de cámaras trampa presenta varias ventajas pues, además de proveer fotografías y videos que permiten probar fehacientemente los hallazgos, facilitan el estudio de especies elusivas o raras con un esfuerzo relativa- mente pequeño (Rowcliffe et al., 2008), así como describir diversos aspectos ecológicos como el uso de hábitats (Head et al., 2012), patrones de actividad (Michalski & Norris, 2011; Rowcliffe et al., 2014) y abundancia relativa (Jenks et al., 2011), entre otros.
En este trabajo presentamos los resultados de una evaluación de mamíferos de hábitos terrestres realizada con cámaras trampa en la zona de uso turístico de la RNAM entre mayo y octubre (temporada de bajas precipitaciones) del 2021. El objetivo principal de dicha evaluación fue obtener una lista de especies que pueda servir de base para el monitoreo de la diversidad de la reserva. De forma adicional, los datos obtenidos fueron utilizados para generar patrones de acti- vidad horaria.
MATERIALES Y MÉTODOS
La zona de uso turístico visitada se encuentra en el sureste de la RNAM. La reserva se ubica en la provincia de Maynas, región de Loreto, a 20 km al suroeste de Iquitos que, con cerca de medio mil- lón de habitantes, es la ciudad más poblada de la Amazonía peruana (Fig. 1). Esta área protegida tiene 58069,9 ha de extensión y es una de las dos únicas en el Perú que conservan ecosistemas de varillales (bosques sobre substrato de arena blanca) y de bosques inundables de aguas negras. El clima en esta región es húmedo y cálido, con una precipitación anual promedio de 3000 mm y una temperatura promedio de 26°C. Aunque la variación de la precipitación es baja a lo largo del año, se reconoce un periodo de menor cantidad de lluvia entre mayo y agosto (Marengo, 1998).
La fase de campo de este estudio fue realizada entre el 02 de mayo y el 08 de octubre del 2021. Durante este periodo, entre seis y diez trampas cámara (Bushnell HD) con iluminación infrarroja estuvieron funcionando al mismo tiempo. Ese número varió debido a que no todas las cámaras se encontraron funcionales o disponibles en el mismo momento. Se colocó las cámaras atándolas a troncos de árboles a 30-40 cm del suelo, apuntando horizontalmente (de forma paralela al suelo) a caminos de animales o a zonas despejadas dentro del bosque. La distancia mínima entre cámaras fue de 250 m, cubriéndose un área de 956000 m2 estimados con el método del polígono convexo mínimo (Meek et al. 2014). Se colocó una sola cámara en cada estación de muestreo, con un total de 10 estaciones de mues- treo. Siendo el objetivo de este estudio realizar un inventario de especies, el área total evaluada y la distancia entre cámaras tendrían poco impacto sobre el resultado (Tobler et al. 2009). Las cámaras fueron programadas para tomar una foto y un video de 10 segundos por evento, con un periodo de pausa de cinco minutos luego de cada evento registrado, y mostrando la fecha y hora en cada fotografía y video. Las ramas y hojas existentes hasta 2 metros frente a las cámaras fueron retiradas para evitar que su movimiento pudiera accionar la toma de imágenes. No se colocó atrayentes o cebos. Los puntos en los que las cámaras fueron colocadas pueden apreciarse en el mapa de la Figura 1. Para confeccionar la lista de especies registradas, se siguió la taxonomía y los nombres comunes presentados por Sánchez-Vendizú, et al. (2021) para los marsupiales y roedores pequeños y por Díaz et al. (2021) para los mamíferos mayores.
La riqueza real de especies fue estimada utilizando el programa EstimateS 9.1.0 (Colwell, 2013), con los índices de riqueza ICE, Chao2 y Jacknife2, que utilizan los datos de incidencia de especies en la muestra (Gotelli & Colwell, 2001). Se tomó cada día de muestreo como una mues- tra (Tobler et al. 2008), teniendo 159 muestras (días) en total. Con el fin de estimar la confiabilidad de los resultados, se graficó una curva de acumulación de especies por rarefacción y curvas del comportamiento de los índices de riqueza al variar el tamaño de la muestra (Colwell et al., 2012), con el uso del programa Excel y los resultados arrojados por EstimateS. Un comportamiento asintótico de las curvas es indicativo de la confiabilidad de los resultados de riqueza real (Gotelli & Colwell, 2001). Este proceso se llevó a cabo considerando todas las especies registradas y, aparte, excluyendo a las especies de comporta- miento básicamente arborícola.
Para cada especie, se calculó dos indicadores de la facilidad para registrar ejemplares: el esfuerzo necesario para registrar el primer evento y la frecuencia de captura. El esfuerzo realizado para lograr el primer registro de cada especie fue expresado en trampas cámara/día (TCD = la suma de cámaras trampa en funcionamiento por cada día). La frecuencia de captura fue calculada como el número de registros independientes por cada 1000 TCD (Tobler et al., 2008). Se consideró registros independientes a los siguientes casos: a) registros consecutivos de diferentes especies, b) registros consecutivos de la misma especie, tomados en diferentes periodos de una hora y c) registros de la misma especie pero no consecutivos, es decir separados por un registro de otra especie. Debido a que no pudo diferenciarse individuos dentro de cada especie, se consideró como un único registro a todas las fotografías de una misma especie tomadas por la misma cámara en el mismo periodo de una hora (Norris et al., 2010). Los registros fueron clasificados como diurnos, nocturnos o crepusculares, según el momento en el que fueron realizados. Se definió el crepúsculo para cada día como el periodo entre el amanecer astronómico y la hora de salida del sol (crepúsculo matutino o alba), y entre la hora de puesta del sol y el atardecer astronómico (crepúsculo de la tarde), utilizando el programa Moonrise 3.5 (Sidell, 2002).
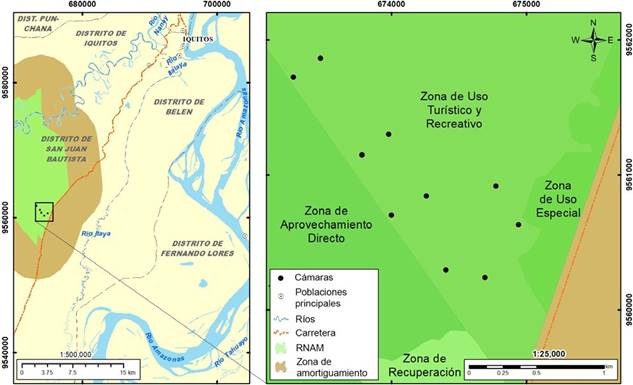
Figura 1 Mapa de ubicación del área de estudio. Los círculos muestran los lugares en los que se colocó trampas cámara. La Reserva Nacional Allpahuayo-Mishana aparece en verde, su Zona de Amortiguamiento en marrón.
Según Lashley et al. (2018), la descripción de un patrón de actividad es confiable cuando se tiene un número de registros cercano o mayor a 100. Siguiendo esta idea, se graficó el horario de actividad de las especies con más de 100 registros utilizando el paquete Overlap (Meredith y Ridout, 2014) con el software R y, de manera referencial, se produjo gráficos para las especies con más de 10 registros independientes utilizando el número de registros obtenidos por cada periodo de una hora (Rowcliffe et al., 2014) con el programa Excel. Las especies con 10 registros o menos no fueron tomadas en cuenta en este proceso por considerarse que tal número es insuficiente (Leuchtenberger et al., 2018).
RESULTADOS
Un esfuerzo de muestreo de 1312 tram- pas cámara/día (TCD) fue llevado a cabo, obteniéndose el registro de 656 eventos indepen- dientes de mamíferos identificados al menos a nivel de género. Un total de 19 especies/géneros de mamíferos fueron registradas (Tabla 1 y Fig.2), incluyendo 18 silvestres y una de origen domés- tico. Por Ordenes, las especies registradas fueron dos marsupiales del Orden Didelphimorphia, cuatro Carnivora, cinco Rodentia, dos Pilosa, dos Cingulata, dos Artiodactyla y un Primate. Fotografías de roedores y marsupiales menores también fueron obtenidas pero no fueron toma- das en cuenta para el presente estudio al no podérseles identificar a nivel de género o especie. La única especie doméstica observada fue el gato doméstico Felis catus, con un solo registro. Dos de los mamíferos registrados tienen costumbres básicamente arborícolas y no suelen ser observa- dos en suelo: el mono ardilla Saimiri sciureus y las ardillas de vientre rojo Hadrosciurus sp., mien- tras que los 17 restantes poseen un componente de actividad terrestre importante.
Tabla 1 Mamíferos registrados por fototrampeo en la RNAM
| Nombre científico | Nombre común | N | f | CTD al 1er evento |
|---|---|---|---|---|
| Didelphimorphia | ||||
| Didelphis marsupialis | Zarigüeya orejinegra | 12 | 9.1 | 60 |
| Metachirus myosuros Cingulata | Rata marsupial | 39 | 29.7 | 20 |
| Dasypus sp. | Armadillo | 35 | 26.7 | 60 |
| Cabassous unicinctus Pilosa | Armadillo de cola desnuda | 1 | 0.8 | 1137 |
| Tamandua tetradactyla | Oso hormiguero amazónico | 2 | 1.5 | 20 |
| Myrmecophaga tridactyla Primates | Oso hormiguero | 2 | 1.5 | 221 |
| Saimiri sciureus Rodentia | Mono ardilla | 2 | 1.5 | 644 |
| Hadrosciurus sp. | Ardilla roja | 19 | 14.5 | 120 |
| Proechimys sp. | Rata espinosa | 38 | 29.0 | 90 |
| Dasyprocta fuliginosa | Añuje | 387 | 295.0 | 10 |
| Myoprocta pratti | Punchana | 1 | 0.8 | 100 |
| Cuniculus paca Carnivora | Majás | 45 | 34.3 | 20 |
| Leopardus pardalis | Tigrillo | 4 | 3.0 | 130 |
| Felis catus | Gato doméstico | 1 | 0.8 | 889 |
| Eira barbara | Manco | 46 | 35.1 | 80 |
| Nasua nasua | Coatí de cola anillada | 4 | 3.0 | 812 |
| Procyon cancrivorus Cetartiodactyla | Mapache cangrejero | 4 | 3.0 | 628 |
| Dicotyles tajacu | Sajino | 8 | 6.1 | 40 |
| Mazama nemorivaga | Venado cenizo | 5 | 3.8 | 90 |
N=número de eventos independientes, f=frecuencia de registro por cada 1000 trampas cámara/día, TCD=trampas cámara/día.

Figura 2 Algunos mamíferos medianos o grandes fotografiados con cámaras trampa en la Reserva Nacional Allpahuayo Mishana. A) Myoprocta pratti, B) Dasyprocta fuliginosa, C) Mazama nemorivaga, D) Myrmecophaga tridactyla, E) Eira barbara, F) Dicotyles tajacu.
Considerando la totalidad de especies registradas, los índices ICE, Chao2 y Jacknife2 estimaron la riqueza de especies en 21.25, 20.11 y 22.33, respectivamente. Al excluir las especies arboríco- las, estos valores fueron de 19.60, 18.30 y 20.21. La curva de acumulación de especies y las curvas de los índices de riqueza mostraron un comportamiento asintótico en ambos casos (Figura 3).
El esfuerzo necesario para registrar el primer evento varió entre 10 TCD para el añuje y 1137 TCD para armadillo de cola desnuda. La frecuencia de captura más elevada correspondió al añuje, con 295.0 registros/1000 TCD (con 387 registros independientes), mientras que la más baja fue de 0.8 registros/1000 TCD para el armadillo de cola desnuda, la punchana y el gato doméstico, que fueron registrados una sola vez cada uno (Tabla 1).
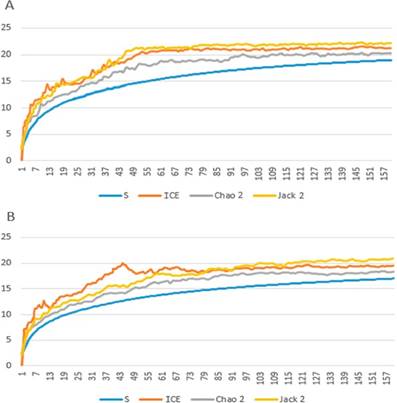
Figura 3 Curvas de acumulación de especies (S) y de los índices de riqueza ICE, Chao 2 y Jacknife 2. Se muestra el número estimado de especies (vertical) y el tamaño muestral, expresado en días de muestreo (horizontal). A) considerando todas las especies registradas, B) exceptuando las especies de costumbres básicamente arborícolas.
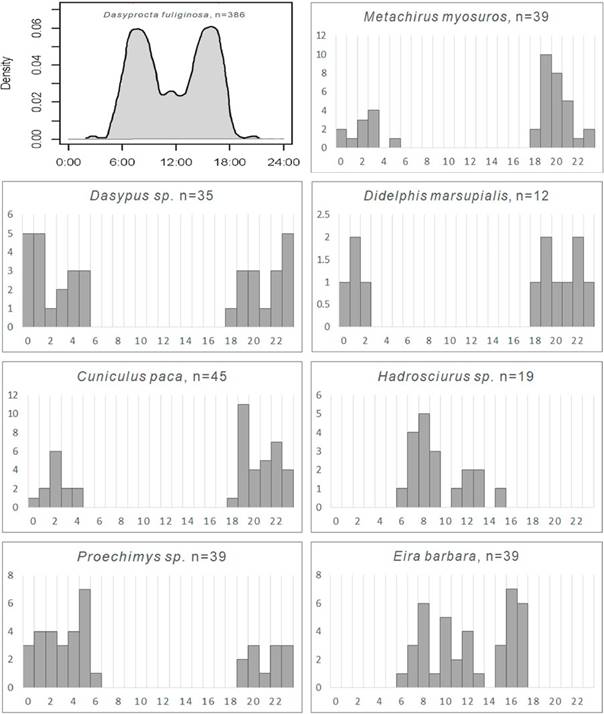
Figura 3 Patrones de actividad de mamíferos registrados por fototrampeo en la RNAM. Horizontalmente, se muestra los intervalos horarios a lo largo de un periodo de un día. Para Dasyprocta fuliginosa se presenta un gráfico realizado con el programa Overlap, indicando verticalmente la fracción de registros correspondientes a cada momento del día. Para las demás especies, con menos de 100 registros, se presenta gráficos preparados en Excel, mostrando verticalmente, se indica el número de registros. El número total de registros para cada especie se muestra luego de cada nombre científico.
Tabla 2 Porcentaje de registros tomados durante el día, la noche y el crepúsculo, para cada especie registrada con trampas cámara en la RNAM.
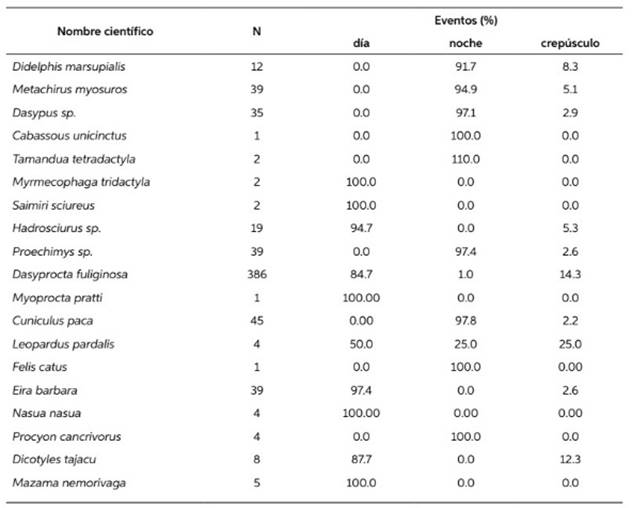
Se consiguió más de 10 registros y se graficó el horario de actividad para ocho de las especies registradas. Cinco (D. marsupialis, M. myosuros, Dasypus sp. C. paca y Proechimys sp.) mostraron actividad principalmente nocturna, con cierta actividad crepuscular, mientras tres (Hadrosciurus sp., E. barbara y D. fuliginosa) fueron principalmente diurnas (Figura 4). El porcentaje de registros conseguidos para cada especie de noche, de día y durante el crepúsculo se muestra en la Tabla 2.
DISCUSIÓN
Dos publicaciones anteriores ofrecen listas de mamíferos terrestres para la RNAM: Oversluijs (2003) menciona 34 especies sujetas a caza registradas en la Zona Reservada Allpahuayo- Mishana, que daría luego lugar a la RNAM. De dicha lista, 16 especies tienen costumbres básica- mente terrestres, y 10 de ellas fueron registradas únicamente de forma indirecta, por huellas. Por otro lado, Hice & Velazco (2012) proporcionan una lista de 13 especies de marsupiales y 24 de roedores capturadas en la reserva, además de mamíferos de otros órdenes de presencia probada o probable en el área.
En el presente trabajo, tres identificaciones fueron hechas sólo a nivel de género: a) los armadillos del género Dasypus, de los cuales se ha registrado anteriormente dos especies en la RNAM (D. novemcinctus y D. pastasae), se diferencian básicamente por la presencia o ausencia de espolones en las rodillas, que son muy difíciles de observar en fotografías; sólo D. novemcinctus ha sido colectado en la RNAM (Hice & Velazco, 2012) mientras que D. pastasae ha sido registrado por huellas (Oversluijs, 2003) y su presencia en la reserva debería ser verificada. b) las ardillas rojas del género Hadrosciurus son difíciles de diferenciar sin capturar individuos (Jessen et al., 2013); sólo H. pyrrhinus ha sido registrada en la RNAM (Abreú et al., 2020), aunque fue inicialmente reportada como H. igniventris (Hice & Velazco, 2012), pero más de una especie podría estar real- mente presente en el área. c) las ratas espinosas del género Proechimys son todas muy parecidas; en la RNAM se ha registrado cuatro especies de este género (Hice & Velazco, 2012).
El hormiguero gigante no había sido registrado antes de forma directa o por huellas en la RNAM. El único indicio de su presencia histórica era una piel encontrada en un establecimiento de hospedaje y se consideró que la especie podía estar localmente extinta debido a la presión de caza (Hice & Velazco, 2012). Este animal suele ser raro donde se le encuentra y es considerado una especie amenazada, en la categoría Vulnerable (VU), tanto a nivel nacional (Cossios, 2018) como global (Miranda et al., 2014). Las fotografías obte- nidas aquí son los primeros registros directos de hormiguero gigante en la RNAM.
Las curvas de especies observadas y de esti- madores de riqueza sugieren que el número de especies registradas en nuestra evaluación se encuentra cerca del número real en el área evaluada, con una diferencia de hasta tres especies. Sin embargo, varios mamíferos registrados por Oversluijs (2003) e Hice & Velazco (2012) no fue- ron observados en esta evaluación, en especial mamíferos grandes como el armadillo gigante Priodontes maximus, tapir Tapirus terrestris, venado colorado Mazama americana, huangana Tayassu pecari, y carnívoros como el otorongo Panthera onca, perro de monte Speothos venaticus, nutria Lontra longicaudis, y hurón grande Galictis vittata. La ausencia de estas especies en la presente evaluación puede deberse a que el área evaluada tuvo una extensión pequeña, menor a 1 km2 y al ámbito hogareño de ciertos mamíferos grandes como el otorongo y el tapir, pero también puede estar influenciada por la cercanía a una gran ciudad, la presencia de una carretera y la ocurrencia de diversas activida- des humanas, incluyendo la cacería en zonas aledañas y el turismo. En zonas tropicales, la densidad poblacional de mamíferos grandes declina con la proximidad a carreteras y otras infraestructuras, hasta a cientos de metros de distancia (Benitez-López et al., 2010), y se ha estimado que la abundancia en zonas con cacería es menor en un 83%, en promedio, con respecto a áreas no disturbadas (Benitez-López et al., 2017). Por su parte, la actividad turística puede también reducir localmente la diversidad de mamífe- ros, así como la abundancia de ciertas especies (Hidinger, 1996; Grossberg et al., 2003; Cunha, 2010; Geffroy et al., 2015).
La actividad horaria de algunas especies también puede verse modificada por actividades antrópicas. La caza y el turismo pueden causar una reducción de la actividad en los momentos con mayor presencia humana, generalmente de día, y aumentarla cuando dicha presencia se reduce (Keuling et al., 2008; Little et al., 2016; Marchand et al., 2014; Ouboter et al., 2021). Asimismo, las modificaciones de hábitat pueden llevar a cambios en la actividad horaria, como sucede con el armadillo de nueve bandas, que aumenta notablemente su actividad diurna en bosques reducidos a parches (Norris et al., 2010).
Nuestros resultados concuerdan con los patrones de actividad anteriormente reportados para la zarigüeya orejinegra, la rata marsupial, los armadillos Dasypus, las ardillas rojas, las ratas espinosas, el añuje, el majás y el manco (Albanesi et al., 2016; Fabre et al., 2016; Wilson et al., 2016; Huck et al., 2017; Leuchtenberger et al., 2018; Mosquera-Guerra et al., 2018; Cossios & Ricra, 2019), aunque deben ser tomados con cuidado debido al bajo número de registros obtenido.
CONCLUSIONES
El muestreo con cámaras trampa permitió estimar de forma confiable la riqueza de mamí- feros terrestres en la zona turística de la RNAM, excluyendo pequeños roedores y marsupiales, con una riqueza observada de 17 especies y una riqueza esperada de entre 18 y 20. Asimismo, se consiguió registrar una especie amenazada y de gran tamaño que se creía extirpada del área, el hormiguero gigante, confirmando su presencia actual en la RNAM.

