











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
La región Loreto tiene una importante diversidad herpetológica con 414 especies de anfibios y reptiles (Aguilar et al., 2021a, 2021b) haciendo que sea un lugar estratégico para la conservación de recursos genéticos (Brooks et al., 2002). Esta importante diversidad es impactada cada vez más por diferentes tipos de presiones antrópicas, tales como: (i) El crecimiento urbano, intensificando el impacto en el deterioro y perdida de los hábitats (Laurance et al., 2009, 2002). (ii) el desarrollo de carreteras, acelerando los procesos de deforestación al hacer accesibles zonas antes aisladas lo que favorece el crecimiento periurbano y el aumento de las superficies agropecuarias (de Souza & Lemos, 2014; Fraser, 2014; Morton et al., 2006; Peres & Schneider, 2012); (iii) Estas actividades generan contaminación en los ecosistemas acuáticos y semiacuáticos (Lebreton et al., 2017; Livesley et al., 2016; Williams et al., 1997) favoreciendo al desarrollo de nuevos patógenos o parásitos que afectan directamente a estas especies, ocasionando disminuciones poblacionales y poniendo en peligro a las especies localmente.
Ante la pérdida masiva de biodiversidad, es necesario incrementar los conocimientos ecológicos, y en particular de los parches de bosques en zonas periurbanas como una estrategia de conservación y mantenimiento de la fauna nativa localmente (Chambers & Hebert, 2016; García-Grajales et al., 2018). Los parches de bosques periurbanos no solo favorecen la conservación de las especies presentes en la zona, si no también mejoran la calidad de vida de los ciudadanos a través de los servicios ecosistémicos que proveen como la reducción de emisiones de carbono y el mejoramiento de la calidad de agua, suelo y aire (Conway et al., 2019; Escobedo et al., 2011; Livesley et al., 2016). Muchos lugares aún siguen sin ser debidamente explorados o evaluados, por lo que no se cuenta un conocimiento sobre su biodiversidad y estado de conservación de las especies. Tal es el caso del Centro de Investigaciones Fernando Alcántara Bocanegra (CIFAB) que a pesar de encontrarse bajo la jurisdicción del IIAP no se tiene conocimiento de su diversidad faunística o florística por lo que se vuelve difícil sustentar el mantenimiento de este tipo de bosque dentro de una zona periurbana. Es por ello que el presente trabajo evalúa la diversidad herpetofaunistica del CIFAB, para comprender los patrones de distribución de especies poco comunes en parches periurbanos y poder formar una línea base de información para toma de decisiones en conservación.
MATERIALES Y MÉTODOS
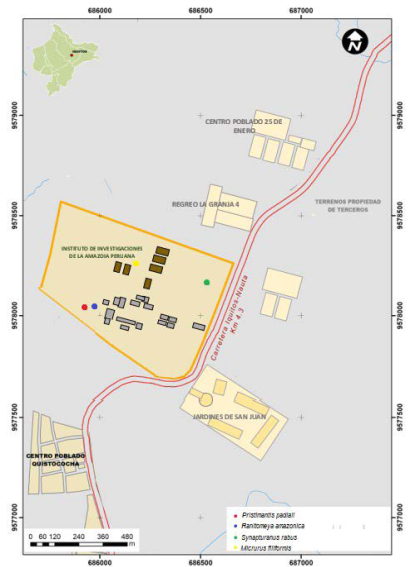
Lugar de estudio: El Centro de Investigaciones Fernando Alcántara Bocanegra (CIFAB), del Instituto de Investigaciones de la Amazonía Peruana (IIAP) ubicado en la carretera Iquitos Nauta kilómetro 4.5, distrito de San Juan, Provincia de Maynas, Departamento de Loreto (03°48´52.4′′ S; 73°19´11.5′′ W) (Figura 1). En sus instalaciones podemos encontrar además de los laboratorios y piscigranjas, pequeños parches de bosques de arena blanca con vegetación característica (García-Villacorta et al., 2003) y aguajales
Muestreo: Se realizaron muestreos de anfibios y reptiles en los meses de junio y setiembre de 2021, con el objetivo la obtener material fresco de tejido para la caracterización a través de código de barras genético. Se realizaron búsquedas libres en los bosques del CIFAB entre las 09:00 horas y las 13:00 horas y se colocaron trampas pitfall durante 7 días del mes de junio. Todos los individuos de anfibios y reptiles capturados fueron fotografiados y sacrificados con benzocaína 7.5% (50 mg/g). Previo al proceso de fijación en formalina al 10%, se extrajo una muestra de tejido muscular para ser almacenada en alcohol 96° y usada en posteriores estudios de ADN. Una vez realizado estos procedimientos, los ejemplares fueron preservados en alcohol al 70% y fueron depositados en la Colección Referencial de Biodiversidad del Instituto de Investigaciones de la Amazonía Peruana (CRBIIAP). La identificación de los especímenes se realizó siguiendo las descripciones originales y literatura pertinente para cada especie (Fouquet et al., 2021; Günter, 1859; Moravec et al., 2010; Schulte, 1999).
RESULTADO
En los muestreos realizados durante los meses de junio y setiembre se registraron un total de 60 individuos pertenecientes a 17 familias, 30 géneros y 40 especies entre anfibios y reptiles siendo los anfibios el clado más abundante con 42 individuos incluidas en ocho familias, 16 géneros y 24 especies, mientras que los reptiles con 18 individuos incluidos en nueve familias, 14 géneros y 16 especies (Tabla 1).
Tabla 1 Lista de especies de anfibios y reptiles registradas en los alrededores de las instalaciones del Centro de Investigación Fernando Alcántara Bocanegra.
| Orden | Familia | Nombre Científico | Código Colector | Código Depósito |
|---|---|---|---|---|
| Anura | Aromobatidae | Allobates femoralis | EJRA1006 | CRBIIAP-2856 |
| Anura | Aromobatidae | Allobates insperatus | EJRA1007 | CRBIIAP-2857 |
| Anura | Bufonidae | Rhinella marina | EJRA1016 | CRBIIAP-2858 |
| Anura | Dendrobatidae | Ameerega hahneli | EJRA1010 | CRBIIAP-2860 |
| Anura | Dendrobatidae | Ranitomeya amazonica | EJRA1053 | CRBIIAP-2855 |
| Anura | Dendrobatidae | Ranitomeya reticulata | EJRA1011 | CRBIIAP-2861 |
| Anura | Hylidae | Boana cinerascens | EJRA1044 | CRBIIAP-2862 |
| Anura | Hylidae | Boana lanciformis | EJRA1036 | CRBIIAP-2863 |
| Anura | Hylidae | Boana punctata | EJRA1040 | CRBIIAP-2864 |
| Anura | Hylidae | Dendropsophus brevifrons | EJRA1045, 1046 | CRBIIAP-2865, 2866 |
| Anura | Hylidae | Dendropsophus leali | EJRA1037, 1039, 1040, 1045 | CRBIIAP-2867, 2868, 2869, 2870 |
| Anura | Hylidae | Dendropsophus triangulum | EJRA1032, 1034, 1035, 1041 | CRBIIAP-2871, 2872, 2873, 2874 |
| Anura | Hylidae | Osteocephalus deridens | EJRA1031 | CRBIIAP-2875 |
| Anura | Hylidae | Scinax ruber | EJRA1022, 1024 | CRBIIAP-2876,2877 |
| Anura | Hylidae | Sphaenorhynchus dorisae | EJRA1019, 1020 | CRBIIAP-2878,2879 |
| Anura | Hylidae | Sphaenorhynchus lacteus | EJRA1017 | CRBIIAP-2880 |
| Anura | Leptodactylidae | Adenomera andreae | EJRA1002,1003, 1011, 1014, 1015 | CRBIIAP-2881, 2882, 2883, 2884, 2885 |
| Anura | Leptodactylidae | Leptodactylus leptodactyloides | EJRA1018, 1043, 1047 | CRBIIAP-2886, 2887, 2888 |
| Anura | Microhylidae | Chiasmocleis bassleri | EJRA1000, 1001, 1051 | CRBIIAP-2889, 2890, 2891 |
| Anura | Microhylidae | Chiasmocleis ventrimaculata | EJRA1027, 1050 | CRBIIAP-2892, 2893 |
| Anura | Microhylidae | Synapturanus sp. "nanay" | EJRA1026 | CRBIIAP-2854 |
| Anura | Strabomantidae | Noblella myrmecoides | EJRA1009 | CRBIIAP-2859 |
| Anura | Strabomantidae | Pristimantis padiali | EJRA1008 | CRBIIAP-2853 |
| Anura | Plethodontidae | Bolitoglossa altamazonica | EJRA1030 | CRBIIAP-2894 |
| Sauria | Boidae | Corallus hortulana | AQUA5 | CRBIIAP-2895 |
| Sauria | Colubridae | Erythrolamprus reginae | AQUA4 | CRBIIAP-2896 |
| Sauria | Colubridae | Helicops angulatus | AQUA1,EJRA1023 | CRBIIAP-2897, 2898 |
| Sauria | Colubridae | Hydrops martii | AQUA3 | CRBIIAP-2899 |
| Sauria | Colubridae | Imantodes cenchoa | EJRA1025 | CRBIIAP-2900 |
| Sauria | Colubridae | Leptophis ahaetulla | AQUA2 | CRBIIAP-2901 |
| Sauria | Dactyloidae | Anolis fuscoauratus | EJRA1012 | CRBIIAP-2902 |
| Sauria | Dactyloidae | Anolis ortonii | EJRA1153 | CRBIIAP-2903 |
| Sauria | Elapidae | Micrurus filiformis | AQUA7 | CRBIIAP-2852 |
| Sauria | Elapidae | Micrurus surinamensis | AQUA6 | CRBIIAP-2904 |
| Sauria | Gymnophthalmidae | Potamites ecpleopus | EJRA1004,1013 | CRBIIAP-2905, 2906 |
| Sauria | Iguanidae | Iguana iguana | EJRA1141 | CRBIIAP-2907 |
| Sauria | Sphaerodactylidae | Gonatodes humeralis | EJRA1029 | CRBIIAP-2908 |
| Sauria | Sphaerodactylidae | Pseudogonatodes guianensis | EJRA1005 | CRBIIAP-2909 |
| Sauria | Teiidae | Dracaena guianensis | EJRA1049 | CRBIIAP-2910 |
| Sauria | Typhlopidae | Amerotyphlops minuisquamus | AQUA8 | CRBIIAP-2911 |
Dentro de esos registros reportamos tres especies raras de anfibios: Pristimantis padiali (Moravec, Lehr, Pérez-Peña, López, Gagliardi-Urrutia, and Arista-Tuanama, 2010) (Strabomantidae) (CRBIIAP-2853), Ranitomeya amazonica (Schulte, 1999) (Dendrobatidae) (CRBIIAP-2855), Synapturanus sp. “nanay” (Fouquet et al., 2021) (Microhylidae) (CRBIIAP-2854) y un ofidio Micrurus filiformis (Günter, 1859) (Elapidae) (CRBIIAP-2852) (Figura 2), en un pequeño parche de bosque de arena blanca rodeado de piscigranjas utilizadas para la cría de peces para el consumo humano.

Figura 2 Especies reportadas en la zona de estudio: (A y B) Pristimantis padiali, (C y D) Ranitomeya amazonica, (E y F), Synapturanus sp. “nanay”, (G y H) Micrurus filiformis.
Nuestros registros añaden una décima localidad conocida para la especie P. padiali (Tabla 2). Consideramos la asignación del individuo CRBIIAP-2853 a P. padiali por presentar discos terminales en los dígitos expandidos, con surcos circunferenciales bien definidos, pliegues dorso laterales ausentes; garganta lisa, vientre débilmente areolado; pliegue discoidal presente; membrana timpánica presente, redonda, alrededor de un tercio de la longitud del ojo; anillo timpánico presente; hocico corto, acuminado en vista dorsal, protuberante e inclinado postero-ventralmente en vista lateral; Dedo I más corto que el Dedo II. En vida presenta la coloración dorsal verde brillante con pequeñas manchas blancas dispersas; cabeza lateralmente marrón claro de noche y marrón oscuro de día; flancos de blanco a amarillo con una franja difusa de color marrón oscuro que delimita la coloración dorsal, área axilar e ingle amarillas; garganta blanca a amarilla, vientre amarillo, superficie palmar y plantar café oscuro; iris café rojizo por la noche y café claro por el día (Moravec et al., 2010).
Tabla 2 Registro de las observaciones de las especies Pristimantis padiali, Ranitomeya amazonica, Synapturanus rabus, Synapturanus sp. “Nanay” y Micrurus filiformis hasta la actualidad.
| Especie | País | Localidad | Longitud | Latitud | Altitud | Fuente |
|---|---|---|---|---|---|---|
| Pristimantis padiali | Perú | Mazan | -73.1819 | -3.5342 | 110 | (Moravec et al., 2010) |
| Pristimantis padiali | Perú | Puerto Almendras | -73.3667 | -3.8167 | 120 | (Moravec et al., 2010) |
| Pristimantis padiali | Perú | Km 26 carretera Iquitos-Nauta | -73.4239 | -3.9636 | 135 | (Moravec et al., 2010) |
| Pristimantis padiali | Perú | Km 31 carretera Iquitos-Nauta | -73.4433 | -4.0069 | S/N | (Moravec et al., 2010) |
| Pristimantis padiali | Perú | Zona Reservada Allpahuayo-Mishana | -73.4333 | -3.9500 | 130 | (Moravec et al., 2010) |
| Pristimantis padiali | Perú | Rio Itaya | -73.7167 | -4.2167 | S/N | (Moravec et al., 2010) |
| Pristimantis padiali | Perú | CIFAB | -73.3219 | -3.8179 | S/N | Este estudio |
| Pristimantis padiali | Perú | Reserva Forestal Santa Cruz | -73.1803 | -3.5214 | 116 | (Metcalf et al., 2020) |
| Pristimantis padiali | Colombia | Quebrada Federico- Bajo Putumayo | S/N | S/N | S/N | (Sánchez et al., 2021) |
| Ranitomeya amazonica | Perú | Puerto Almendras | -73.3667 | -3.8167 | 120 | (Schulte & Hauck, 1999) |
| Ranitomeya amazonica | Perú | Reserva Forestal Santa Cruz | -73.1803 | -3.5214 | 116 | (Metcalf et al., 2020) |
| Ranitomeya amazonica | Perú | CIFAB | -73.3218 | -3.8179 | S/N | Este estudio |
| Ranitomeya amazonica | Perú | Km 26 carretera Iquitos-Nauta | -73.4050 | -3.9640 | S/N | (Brown et al., 2011) |
| Ranitomeya amazonica | Perú | Km 41 Carretera Iquitos - Nauta | -73.4620 | -4.0750 | (Brown et al., 2011) | |
| Ranitomeya amazonica | Colombia | Leticia | -69.9396 | -4.1123 | S/N | (Symula et al., 2003) |
| Synapturanus rabus* | Colombia | Rio Vaupés | -69.4667 | 1.0500 | S/N | (Pyburn, 1976) |
| Synapturanus rabus | Perú | Reserva Forestal Santa Cruz | -73.1803 | -3.5214 | 116 | (López-Rojas & Cisneros-Heredia, 2012) |
| Synapturanus rabus | Perú | Reserva Forestal Santa Cruz | -73.1803 | -3.5214 | 116 | (Metcalf et al., 2020) |
| Synapturanus rabus | Ecuador | Orellana | S/N | S/N | S/N | (Wild, 1995) |
| Synapturanus sp. “nanay” | Perú | CIFAB | -73.3198 | -3.8143 | S/N | Este estudio |
| Micrurus filiformis | Brasil | Pará | S/N | S/N | S/N | (Günter, 1859) |
| Micrurus filiformis | Colombia | Carurú | S/N | S/N | S/N | (Silva, 1994) |
| Micrurus filiformis | Perú | Iquitos | S/N | S/N | S/N | (Dixon & Soini, 1977) |
| Micrurus filiformis | Perú | CIFAB | -73.3217 | -3.8165 | S/N | Este estudio |
* Localidad tipo de Synapturanus rabus sensu stricto.
El individuo CRBIIAP-2855 de R. amazonica coincide con lo descrito por Schulte, (1999), quien caracteriza la especie por presentar: la piel dorsal lisa; los discos del dedo de la mano II y IV muy expandidos; el dedo I es más corto que el dedo II; las rayas dorso laterales varían de color rojo a naranja; la "Y" negra definida en la parte dorsal de su cuerpo, punto negro en el hocico; línea labial de color amarillo; franja lateral de color amarillo; superficie de las extremidades posteriores y vientre negro con reticulaciones que varían desde el azul intenso; garganta brillante de color naranja, con los bordes negros irregulares.
La especie Synapturanus sp. “nanay” fue encontrada dentro de una trampa pitfall colocada por los autores detrás de los estanques de piscicultura. Se identificó mediante comparaciones de los caracteres morfológicos del individuo CRBIIAP-2854 con el ejemplar MZUNAP01.834 reportado por (Fouquet et al., 2021), el cual comparte similitud con el denominado como Synapturanus sp. “nanay”. Además, el individuo que reportamos se encuentra dentro de la cuenca del río Nanay, por lo que por distribución consideramos que su determinación corresponde a Synapturanus sp. “nanay”. Este registro representa una segunda localidad reportada para esa especie no descrita en Perú, siendo parte de la diversidad críptica del género Synapturanus en la Amazonía peruana (Fouquet et al., 2021). Además de este nuevo registro geográfico, se registró la presencia de parásitos nemátodos dentro de su tracto digestivo.
Finalmente, se identificó a CRBIIAP-2852 como Micrurus filiformis por la siguiente combinación de caracteres: cuerpo delgado con un diámetro de 28 mm; hocico negro con una banda blanca sobre las escamas interna- sales, prefrontales y los primeros dos pares de supralabiales; banda negra sobre la mayoría de los escudos frontales y parietales; escamas gulares rojas; cuatro pares de infralabiales uniformemente negras; de 17 tríadas corporales; sin tubérculos supra anales (Feitosa et al., 2007).
DISCUSIÓN
Con la creación de la carretera Iquitos - Nauta se pudo observar una explosión en el crecimiento urbano y periurbano de la ciudad, cada vez se encuentran menos paisajes naturales cerca del eje de la carretera provocando que especies de flora y fauna que antes habitaban desaparezcan (Achung, 1994), durante este tiempo de expansión, se realizaron pocos estudios sobre la composición y distribución de especies de anfibios y reptiles de estas zonas (Dixon & Soini, 1977) por lo que no se tiene conocimiento sobre lo que se podría estar perdiendo.
Las especies que reportamos en este estudio se encuentran en diferentes categorías de conservación (MINAM, 2018) y por lo que se sabe tienen características ecológicas particulares lo que las hace requerir ecosistemas también particulares como los que encontramos dentro del CIFAB, la expansión urbana muchas veces no toma en consideración la preservación de estos tipos de bosques y en consecuencia la preservación de las especies que allí habitan, motivo por el cual estudios que toman en consideración las zonas urbanas y periurbanas son vitales para la conservación de la herpetofauna nativa.
Pristimantis padiali pertenece a la familia Strabomantidae y es una especie que se encuentra en bosques primarios de la Amazonía peruana y colombiana. No existen muchos registros de esta especie, pues se considera que utilizan como principal hábitat el dosel del bosque (Moravec et al., 2010). En su descripción en el 2010, uno de los paratipos asignados de la especie provenía del área de Puerto Almendras que es una localidad cercana al CIFAB (Moravec et al., 2010) sin embargo, hasta la fecha no ha vuelto a ser reportada formalmente en las cercanías a Iquitos (Metcalf et al., 2020; Sánchez et al., 2021), Durante más de una década los asentamientos humanos entre las localidades de Puerto Almendras y el CIFAB se han ido incrementando de manera vertiginosa (Achung, 1994; Moschella, 2019), reduciendo y fragmentando el hábitat, produciendo discontinuidades cada vez mayores entre parches de bosques, causando la declinación de sus poblaciones (Young et al., 2001) . El deterioro del hábitat y la disminución de los tamaños de las poblaciones de esta especie hacen que estos registros sean aún más raros, escasos e inesperados para la zona.
Ranitomeya amazonica es una especie de la familia Dendrobatidae conocida de pocas localidades de la Amazonía occidental (como Brasil Colombia, Ecuador y Perú) (Brown et al., cular, y/o patrones de distribución (Fouquet et al. 2011; Metcalf et al., 2020; Schulte, 1999; Symula et al., 2003). Estudios previos reportan a esta especie en pequeños parches de bosques de arena blanca en las cercanías a Iquitos (Brown et al., 2011). Los registros señalan la existencia de una clara diferenciación entre los morfo de arena blanca y de otras localidades (Brown et al., 2011). Esta es una especie amenazada por la destrucción de los ecosistemas de arena blanca para la extracción de arena y madera redonda para construcción, lo que pone en peligro a las poblaciones de R. amazonica debido a la fuerte relación de esta especie con este tipo de hábitat (García-Villacorta et al., 2003; Mori Vargas & Reátegui Sunción, 2012; Panduro Tejada, 2008). Además de las amenazas por degradación de su hábitat, sufren una presión elevada por extracción y comercio ilegal de especímenes para el comercio de especies ornamentales, pudiendo diezmar sus poblaciones (Auliya et al., 2016). Por ello, el mantener saludables las poblaciones relictas de los parches de bosque como del CIFAB puede contribuir significativamente al mantenimiento de la población de esta especie.
Nuestro registro de Synapturanus sp. “nanay”, una especie poco frecuente de registrar por sus hábitos fosoriales (Fouquet et al., 2021) capturada mediante el uso de trampas de caída (Angulo et al., 2006). Lo que muestra la importancia de emplear varios tipos de técnicas de muestreo para optimizar el registro de un mayor número de especies, sobre todo para aquellas que suelen ser difíciles de ubicar por sus requerimientos ecológicos (López-Rojas & Cisneros-Heredia, 2012). Todas las especies de Synapturanus reportadas en Amazonía oeste (Perú y Colombia) parecen constituir entidades taxonómicas crípticas, difícilmente de diferenciar por la similitud de las características morfológicas externas. Solamente son posibles de diferenciar basados en información osteológica, miológica, acústica, mocular, y/o patrones de distribución (Fouquet et al. 2021).
También se registró una especie de serpiente coral, Micrurus filiformis, la cual ha sido reportada una sola vez para la ciudad de Iquitos por Dixon & Soini (1977). Desde entonces hasta el presente estudio no se habían reportado más registros. A pesar de que esta especie tiene una distribución geográfica muy amplia no es una especie común y fácil de registrar, por lo que es sorprendente e inesperado reportar la especie en un parche periurbano como el que se encuentra en las instalaciones del CIFAB (Feitosa et al., 2007).
CONCLUSIÓN
Los ecosistemas que se encuentran dentro del CIFAB mantienen una inesperada diversidad de anfibios y reptiles a pesar de su ubicación. Este carácter inesperado se debe a las importantes presiones antrópicas que la rodea.
El lugar donde se encuentra el CIFAB fue creado en la década de los 70, hace más de 40 años al inicio de la expansión de la ciudad, y con su creación ayudó a la preservación de su biodiversidad, por lo que es de suma urgencia evaluar la biodiversidad y el estado de salud de las poblaciones que alberga el CIFAB y los parches de bosques periurbanos en general. Estos parches pueden jugar un papel importante en la conservación de las especies, pero también para mejorar la calidad de vida de los ciudadanos por los diversos servicios ecosistémicos que pueden brindar.

