










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkArnaldoa
Print version ISSN 1815-8242On-line version ISSN 2413-3299
Arnaldoa vol.25 no.2 Trujillo mayo/ago. 2018
http://dx.doi.org/http://doi.org/10.22497/arnaldoa.252.25217
ARTÍCULOS ORIGINALES
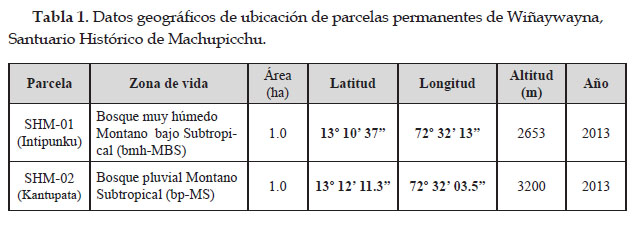
Dinámica, biomasa aérea y variables poblacionales de dos parcelas permanentes en bosques montanos de Wiñaywayna, Santuario Histórico de Machupicchu, Cusco, Perú
Dynamics, Aerial Biomass and Population Variables of two Permanent Plots in Montane Forests of Wiñaywayna, Machupicchu Historic Sanctuary, Cusco, Peru
Lucero E. Alfaro Curitumay, Greta M. Paiva Prado & Heldy Yiyi Espinoza Carrasco1,a, Abel Monteagudo Mendoza2, Wilfredo Chávez Huaman3
1 Universidad Nacional de San Antonio Abad del Cusco, PERÚ
2 Universidad Nacional de San Antonio Abad del Cusco, Jardín Botánico de Missouri, Oxapampa, Pasco, PERÚ
3 Instituto de Manejo de Agua y Medio Ambiente, Cusco, PERÚ
a segundo_leiva@hotmail.com/cleivag@upao.edu.pe
Resumen
En los bosques montanos de las inmediaciones de Wiñaywayna dentro del Santuario Histórico de Machupicchu (SHM), se evaluó la dinámica poblacional, biomasa aérea almacenada y las variables poblacionales de árboles con DAP≥10, de dos parcelas permanentes de 1.0 ha cada una, en los sectores de Intipunku (SHM–01) a los 2653 m de altitud y Kantupata (SHM–02) a los 3200 m de altitud, desde setiembre del 2013 a noviembre del 2016 bajo la metodología estandarizada de RAINFOR. Se registraron un total de 69 especies en 1286 individuos; en la parcela SHM–01, 26 familias, 36 géneros y 45 especies en 682 individuos, donde las especies con mayor IVI fueron Meliosma peytonii con 33.50 %, Hieronyma oblonga con 29.23 % y Gordonia fruticosa con 28.43 %. En la parcela SHM–02, se registró a 22 familias, 28 géneros y 31 especies en 604 individuos, donde las especies con mayor IVI fueron Weinmannia crassifolia con 70.82 % que además fue la más abundante y dominante, Ilex andicola con 35.65 % y Hedyosmum angustifolium con 35.09 %. Se registró una diversidad moderadamente alta, de 3.063 y 2.477 según Shannon, 0.93 y 0.87 según Simpson para Intipunku y Kantupata respectivamente con una muy baja similaridad entre ellas por compartir solo 7 especies; lo que se traduce en una composición de especies arbóreas diferente en cada parcela. En SHM–01 se presentó 1.18 % de tasa de mortalidad, 0.79 % de tasa de reclutamiento y 0.99 % de tasa de renovación anual y en SHM–02, 1.21 % de tasa de mortalidad, 2.05 % de tasa de reclutamiento y 1.63 % de tasa de renovación anual, con un desequilibrio entre dichas tasas. La biomasa aérea almacenada fue mayor en Intipunku de 145 ± 2.80 T ha-1, con una productividad de 0.95 Tha-1 año-1, las especies Gordonia fruticosa y Aniba coto aportaron mayor biomasa con 19 Tha-1 cada una; la biomasa en Kantupata fue de 101.06 ± 3.46 Tha-1, con una productividad de 1.18 T ha-1 año-1, la especie Weinmannia crassifolia aportó mayor biomasa de 32 Tha-1.
Palabras clave: RAINFOR, Bosque montano, biomasa arbórea, dinámica, diversidad, composición florística y parcelas permanentes.
Abstract
In the montane forests near Wiñaywayna within the Historic Sanctuary of Machupicchu (SHM), we evaluated the population dynamics, stored aerial biomass and the population variables of trees with DBH≥10, of two permanent plots of 1.0 ha each one, in the sectors of Intipunku (SHM-01) at 2653 m of altitude and Kantupata (SHM-02) at 3200 m of altitude, from September 2013 to November 2016 under the standardized RAINFOR methodology. We registered a total of 69 species in 1286 individuals; in plot SHM-01, 26 families, 36 genera and 45 species in 682 individuals, where the species with higher IVI were Meliosma peytonii with 33.50 %, Hieronyma oblonga with 29.23 % and Gordonia fruticosa with 28.43 %. In plot SHM-02, 22 families, 28 genera and 31 species in 604 individuals, where the species with the highest IVI were Weinmannia crassifolia with 70.82 %, which was also the most abundant and dominant, Ilex andicola with 35.65 % and Hedyosmum angustifolium with 35.09 %. A moderately high diversity was recorded, of 3.063 and 2.477 according to Shannon, 0.93 and 0.87 according to Simpson for Intipunku and Kantupata respectively with a very low similarity among them for sharing only 7 species, which means a different tree species composition in each plot. In SHM-01 there was 1.18 % mortality rate, 0.79 % recruitment rate and 0.99 % annual renewal rate and in SHM-02, 1.21 % mortality rate, 2.05 % recruitment rate and 1.63 % annual renewal rate, with an imbalance between these rates. The stored aerial biomass was higher in Intipunku at 145 ± 2.80 T ha-1, with a net primary productivity of 0.95 Tha-1year-1, the species Gordonia fruticosa and Aniba coto contributed higher biomass with 19 Tha-1 each; the biomass in Kantupata was 101.06 ± 3.46 Tha-1, with a net primary productivity of 1.18 T ha-1 year-1, the Weinmannia crassifolia species contributed greater biomass with 32 Tha-1.
Keywords: RAINFOR, montane forest, tree biomass, dynamics, diversity, floristic composition, permanent plots.
Introducción
Los bosques montanos son ecosistemas ubicados por encima de los 1000 m y se extiende por la ceja de montaña de la vertiente oriental de los andes, pasando a la vertiente occidental por el norte peruano (Weberbauer 1945). Estas áreas son ecosistemas frágiles, pues contienen una diversidad biológica caracterizada por su alto grado de singularidad y rareza (Cuesta et al., 2009), por el endemismo de su diversidad biológica (Gentry, 1992) y por ser fundamentales en la provisión de servicios ecosistémicos, principalmente vinculados al agua, a la regulación climática regional y a la captura y almacenamiento de carbono (Cuesta et al., 2009), sin embargo, su papel en el ciclo y almacenamiento de carbono es aún poco conocido (Girardin et al., 2013; Spracklen & Righelato, 2013), pese a que son importantes sumideros. Estos ecosistemas hoy en día están siendo vulnerados por la degradación antrópica, hechos que los lleva a ser prioritarios en la conservación.
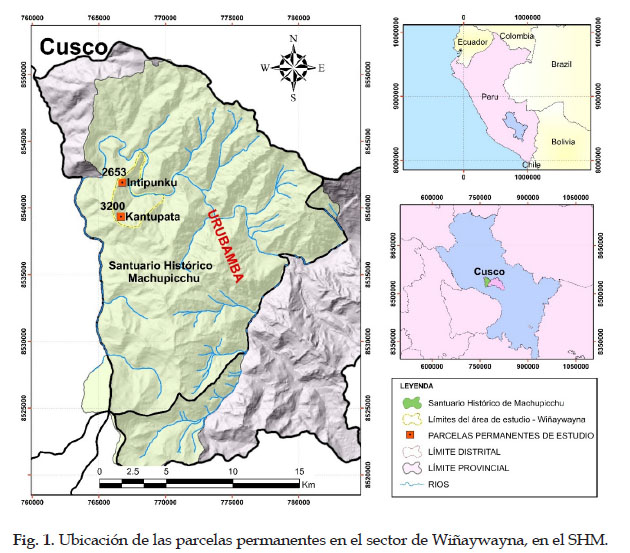
El Santuario Histórico de Machupicchu es una de las áreas naturales protegidas de mayor importancia para el estado peruano y reconocida por la UNESCO, como patrimonio de la humanidad y representa un gran potencial biótico y diversidad de hábitats, sin embargo, el poco conocimiento sobre la dinámica poblacional y almacenamiento de biomasa-carbono de sus bosques montanos, generan vacíos de información, por lo que se evaluó en la presente investigación, la dinámica poblacional, biomasa aérea-carbono almacenado en dos parcelas permanentes preestablecidas en los bosques montanos de los sectores de Intipunku a los 2653 m (SHM–01) y Kantupata a los 3200 m (SHM–02) en Wiñaywayna dentro del Santuario
Histórico de Machupicchu, tomando datos desde el 2013 al 2016, bajo la metodología estandarizada de parcelas permanentes de RAINFOR.
Materiales y métodos
Área de estudio
El presente estudio se llevó a cabo en dos parcelas permanentes instaladas por RAINFOR, que se encuentran ubicadas a través de una gradiente altitudinal, en los sectores de Intipunku a los 2653 m en las coordenadas 13º 10’ 37" Latitud Sur y 72º 32’ 13" Longitud Oeste y en Kantupata a los 3200 m en las coordenadas 13º 12’ 11,3" Latitud Sur y 72º 32’ 03.5" Longitud Oeste, en los bosques montanos de Wiñaywayna dentro del Santuario Historio de Machupicchu, Distrito de Machupicchu, Provincia de Urubamba y Región del Cusco – Perú.
Métodos de campo
La toma de datos se realizó en dos censos, con un intervalo de 3 años en setiembre del 2013 se llevó a cabo el primer censo y noviembre del 2016 el segundo censo, bajo el sistema estandarizado de procedimientos para remedición y establecimiento de parcelas permanentes para bosques montanos establecido por la Red Amazónica de Inventarios Forestales (RAINFOR) 2001, donde se incluyeron todos los individuos de árboles con DAP ≥10 cm.
Análisis de datos
Los datos obtenidos en campo fueron ordenados en una matriz organizada por familias, especies y números de individuos para cada una de las parcelas permanentes, el tratamiento taxonómico siguió el sistema de clasificación APG IV (2016), además se consultó bibliografía correspondientes y páginas web: http://www.tropicos.org, http://fieldmuseum.org, http://tnrs.iplantcollaborative.org.
La composición florística de cada una de las parcelas, se trabajó con todos aquellos arboles encontrados vivos en el último censo, las cuales se ordenaron en una base de datos por familias, géneros y especies, fueron revisados y actualizados con listados en la página web de Trópicos (http://www.tropicos.org/) y Forestplots Database www.foresplots.net.
La dinámica poblacional se analizó mediante3parámetros;tasadereclutamiento (TR = (Ln (No - Nm + Nr) – Ln (No- Nm)) / ∆t), la tasa de mortalidad (TM=((ln(No)– ln(No- Nm))/∆t) y la tasa de renovación de individuos (TRe=(TM+TR)/2), donde No es el número de individuos al comienzo del periodo, Nm es el número de individuos muertos en una determinada área, Nr es el número de individuos reclutados en el segundo periodo y ∆t es la variación del tiempo entre dos periodos de medición (Nebel et al. 2000).
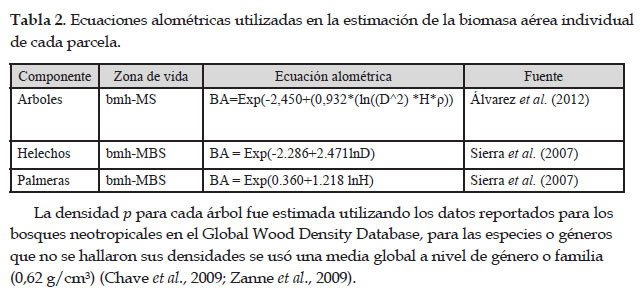
La biomasa aérea para cada árbol se determinó mediante la ecuación alométrica propuesta por Álvarez et al., 2012., según zonas de vida, en función al diámetro a la altura del pecho (d.a.p), altura del árbol y a la gravedad específica de madera o densidad (ρ), la cual se totalizó para toda la parcela como la sumatoria de la biomasa arbórea de todos los arboles por hectarea, con la siguiente formula, (BAt=ΣBAest/ Area) donde, AGB est: Biomasa arbórea sobre el suelo, Área: Tamaño de la parcela en hectárea. (Honorio & Baker 2010). Para los helechos y palmeras, se usó las ecuaciones de Sierra et al., 2007.
Diversidad
La diversidad alfa, se analizó mediante los índices de diversidad de Shannon – Wiener, este Índice combina la información de la riqueza de especies y la equidad, en la diversidad o heterogeneidad (Magurran 2004; Moreno et al., 2006), cuanto mayor sea del número de especies, incrementa la diversidad, así como la uniformidad (Franco, 1989), y se analizó mediante la siguiente formula (H’=Σpi*lnpi), donde H’: Índice de Shannon – Wiener, Ln: Logaritmo natural y pi: Abundancia proporcional de la especie i (ni/Nt). Y mediante el índice de Simpson, este índice es inverso al concepto de equidad de la comunidad ya que toma en consideración las especies con mayor importancia ecológica sin considerar al resto de las especies, y es menos sensible con la riqueza de especies (Krebs, 1978; Magurran 1991; Feinsinger, 2003), se analizó mediante la siguiente ecuación D=∑[ni(ni-1)/N(N-1) ], donde D= Índice de Simpson, ni= Número de individuos de la especie i en la comunidad 1, N= Número total de individuos de la comunidad 1, (Moreno, 2001).
La diversidad beta para comparar comunidades se analizó mediante el Índice de similaridad de Morisita – Horn, la cual calcula la similitud en base a datos cuantitativos y es muy sensible a la abundancia de las especies (Magurran, 1988; Baev & Penev, 1995), por lo que, Wolda (1983) propuso una modificación para este índice, se cálculo con la siguiente ecuación: CMH = 2Σ(ani1x bnj)/(da+db) aNxbN*100% ; da = (Σan1)2/(aNa)2; db = (Σbnj)2/(bNb)2, donde, aN: Número total de individuos en el sitio A, bN: Número total de individuos en el sitio B, ani: Número de individuos de la i-esima especie en el sitio A, bnj: Número de individuos de la i-esima especie en el sitio B.
Importancia ecológica (IVI’s)
La importancia ecológica o IVIs, se analizó como la sumatoria de las variables frecuencia relativa (Fr), Dominancia relativa (Dr) y Abundancia relativa (Ar), analizadas para cada parcela.
Resultados
Composición florística
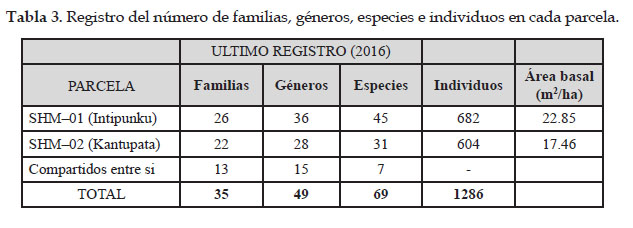
Se evaluaron un total de 1286 individuos de árboles con DAP ≥ 10 cm pertenecientes a dos parcelas permanente de 1.0 – ha de bosque cada una, para el bosque de Intipunku (SHM–01), se evaluó 682 individuos pertenecientes a 26 familias con 36 géneros y 45 especies con las familias Cyatheaceae con 6 especies, Melastomataceae con 5 especies, Rubiaceae con 4 especies y Euphorbiaceae con 3 especies como las más diversas y las familias Sabiaceae con 104 individuos, Melastomataceae y Phyllanthaceae con 73 como las más abundantes.
Para el bosque de Kantupata (SHM–02) se evaluó a 604 individuos pertenecientes a 22 familias con 28 géneros y 31 especies; las familias Lauraceae y Melastomataceae con 3 especies son las más diversas y la familia Cunoniaceae fue la más abundante por presentar 180 individuos por hectárea, seguida de Chloranthaceae con 85 y Aquifoliaceae con 76 individuos.
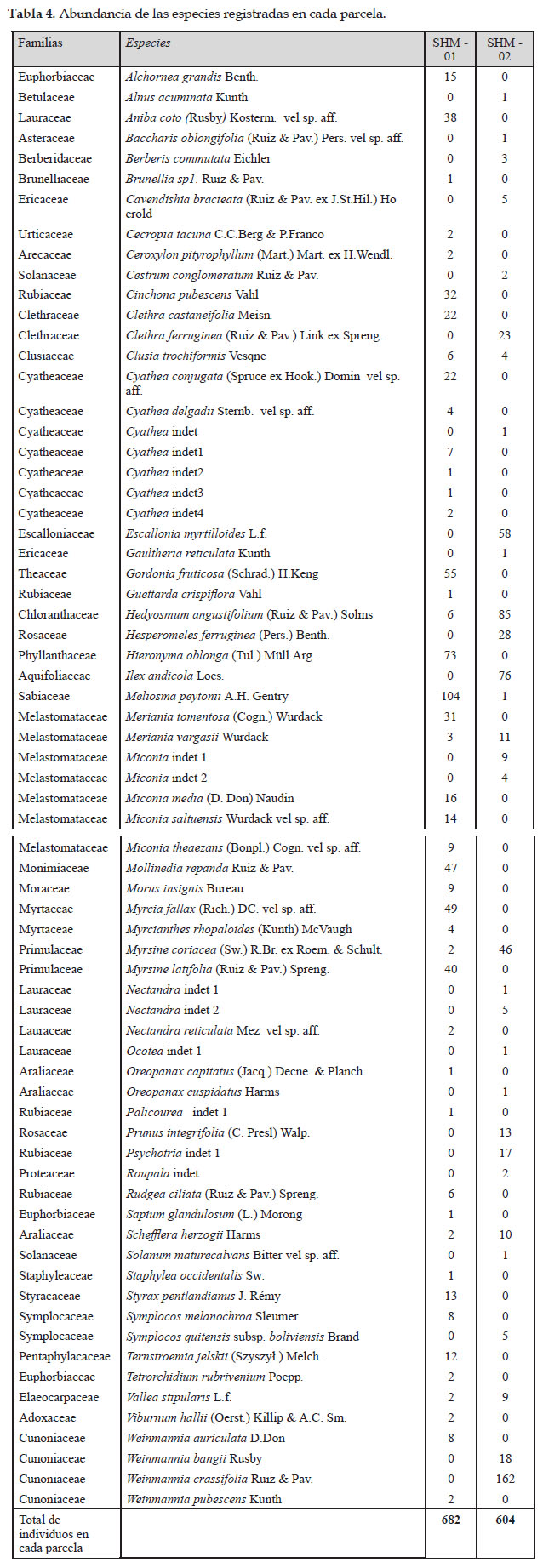
Dinámica poblacional
En la parcela de Intipunku se registró mayor tasa de mortalidad de 1,18 %, respecto a su tasa de reclutamiento que solo fue de 0,79 % y su tasa de recambio que fue de 0,99 %. En la parcela de Kantupata en cambio se registró mayor tasa de reclutamiento de 2,05 % respecto a su tasa de mortalidad que fue de 1.21 %, su tasa de recambio fue de 1,63 %. Se muestra que la parcela SHM–02 registro los valores más altos que la parcela SHM–01, quiere decir el bosque de Kantupata tuvo más dinámica con más reclutamiento y menos mortalidad,
que el bosque de Intipunku que tuvo más mortalidad y menos reclutamiento.
Para ser los primeros resultados de dinámica poblacional en los bosques estudiados dentro del Santuario Histórico de Machupicchu, se muestra que ambas parcelas tienen un relativo desequilibrio entre las tasas de mortalidad y reclutamiento, esto se puede deber a que los bosques son jóvenes o bosques no maduros que están en proceso de crecimiento y recuperación como es el caso del bosque de Intipunku que en algún momento ha sufrido intervención antrópica.
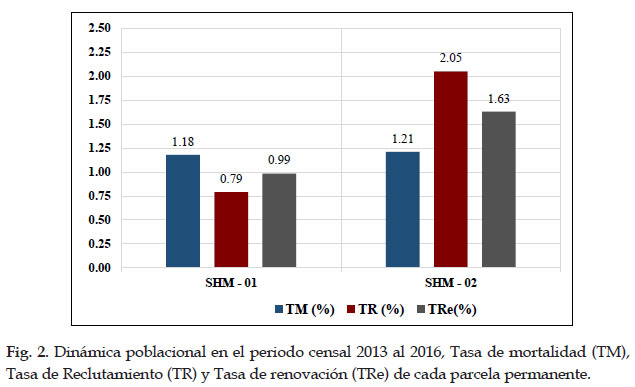
Biomasa aérea almacenada
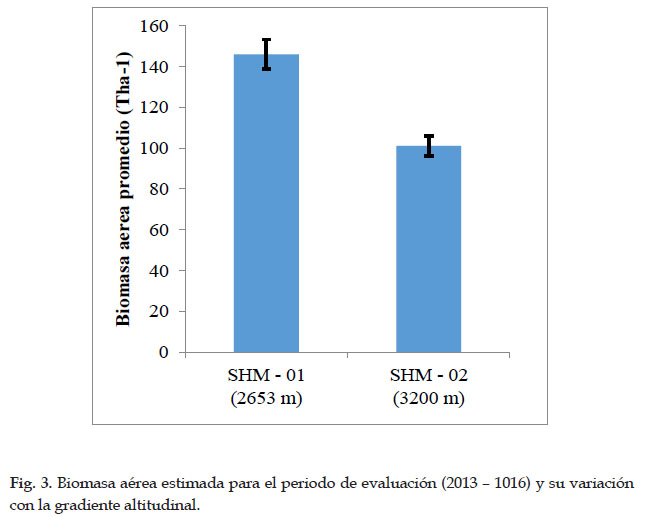
En la parcela SHM–01 del bosque de Intipunku, se registraron un total 682 individuos con DAP ≥ 10 cm, de los que se obtuvo la biomasa aérea total de 145,98± 2,80 T ha-1, en la parcela SHM–02 del bosque de Kantupata se registraron 604 individuos con DAP ≥ 10, lo que representó en 101,06± 3,46 T ha-1 de biomasa aérea total.
Se determinó una relación negativa de gradiente y biomasa, quiere decir que a mayor altitud menor cantidad de biomasa aérea, en la parcela SHM–01 de Intipunku que se encuentra a 2653 m se registró mayor biomasa aérea y carbono acumulado que en la parcela SHM–02 de Kantupata la cual se encuentra a mayor altitud (3200 m), esto se debe al tipo de bosque, a la dinámica que ocurre en ella y a la composición florística diferente en cada una de ellas, pues influyen en el crecimiento diamétrico, altura arbórea y densidad de madera. Esto fue corroborado con el estadístico no paramétrico de U de Mann Whitney, según el cual, el P valor=0.001 al 95% resulto ser menor que 0,05 (P<0,05), por consiguiente, se aceptó la hipótesis alterna que indica que existe una marcada diferencia significativa en las medianas de biomasa aérea almacenada entre ambas parcelas.
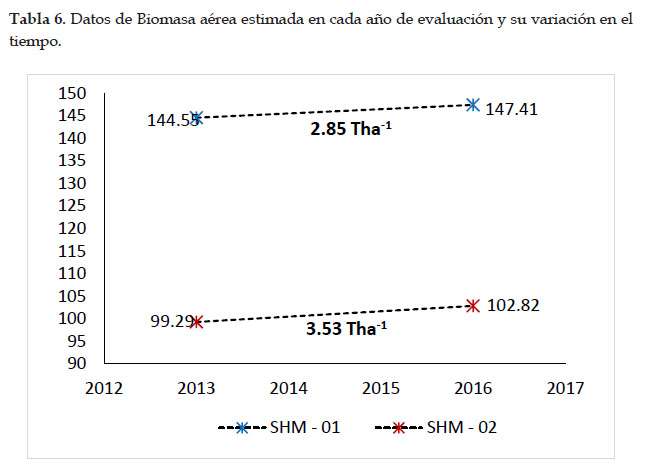
En los 3 años del periodo intercensal (2013 al 2016), se registró un incremento de 2,85 T ha-1 de biomasa aérea en la parcela SHM–01 de Intipunku y un incremento de 3,53 Tha-1 de biomasa aérea en la parcela SHM–02 de Kantupata y fue esta la que presentó mayor variación.
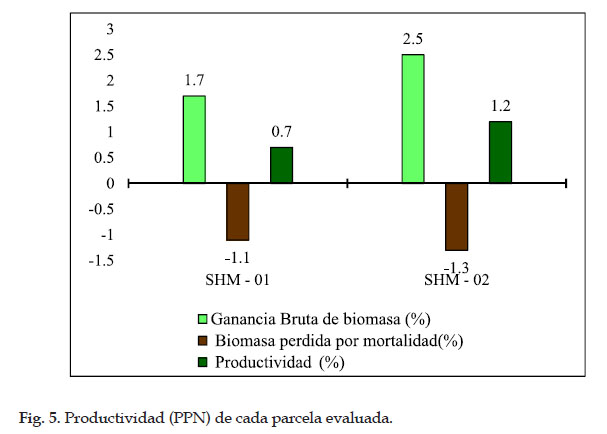
La productividad registró una pérdida de biomasa por mortalidad de 1,1% anual y ganancia bruta (por crecimiento más reclutamiento) de 1,7% anual, que resulta en una productividad de 0,7% en la parcela SHM–01 de Intipunku. Mientras que en la parcela SHM–02 de Kantupata, se registró una pérdida de biomasa por mortalidad de 1.2% al año, una ganancia bruta (por crecimiento más reclutamiento) de 2,4% al año y una productividad de 1,2% al año, siendo esta parcela la que presentó mayor Poductividad.
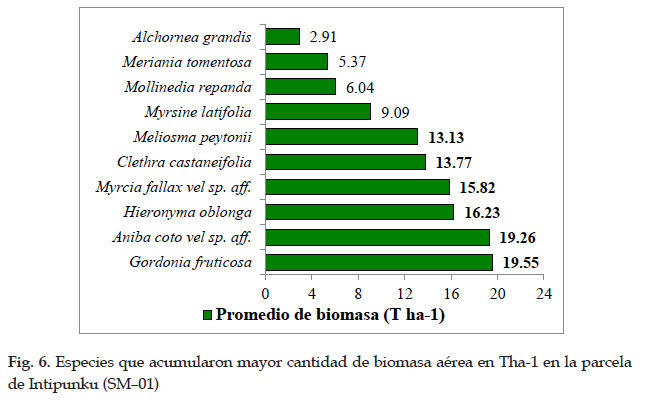
Las especies Gordonia fruticosa (19,55 Tha-1) y Aniba coto vel sp. aff. (19,26 Tha-1), son los que aportaron mayor biomasa aérea, seguido de Hieronyma oblonga (16,23 Tha-1) y Myrcia fallax vel sp. aff. (15, 82 Tha- 1), seguido de Clethra castaneifolia (13,77 Tha-1) con Meliosma peytonii (13,13 Tha-1), estas especies acumularon casi el 80% de biomasa aérea por hectárea en la parcela SHM–01.
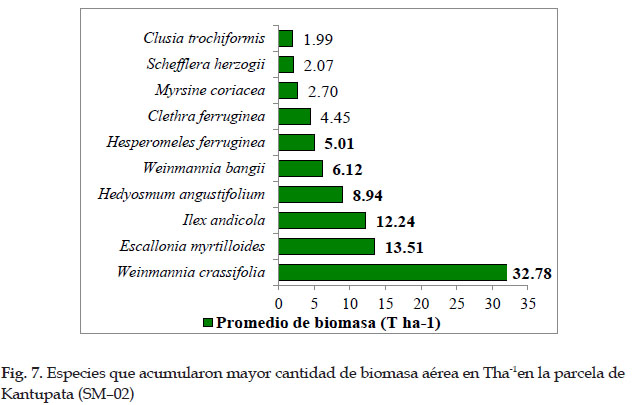
La especie Weinmannia crassifolia, tuvo el mayor aporte de biomasa aérea con 32,78 Tha-1, seguida de Escallonia myrtilloides 13,51 Tha-1, Ilex andicola 12,24 Tha-1, Hedyosmun angustifolium con 8,24 Tha-1, Weinmannia bangii con 6,12 Tha-1, Hesperomeles ferruginea con 5,01 Tha-1, estas especies acumularon mayor biomasa aérea por hectárea representando casi el 89 % del total en la parcela SHM–02.
Diversidad
El índice de Shannon Wiener en base a la riqueza de especies indica una diversidad moderadamente alta de 3,063 para la parcela de Intipunku y de 2,477 para la parcela de Kantupata y el Índice de Simpson en base a la abundancia indica una diversidad alta de 0,93 para la parcela de Intipunku y 0,87 para la parcela de Kantupata, según estos resultados la parcela de Intipunku fue más diversa que la parcela de Kantupata.
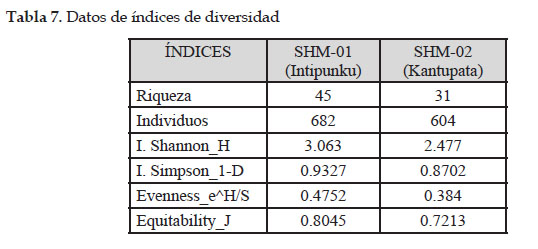
El índice de Equidad de Pielou muestra una distribución proporcional y casi equitativa del número de especies por número de individuos por presentar valores próximos a 1, siendo para la parcela de Intipunku el valor de 0,80 y para la parcela de Kantupata el valor de 0,72.
Según el Índice de Morissita Horm, las parcelas presentan una similitud muy baja de solo 1,97 %, quiere decir que son diferentes y entre sí solo comparten 7 especies, esto se debe a la gradiente altitudinal de 653m que existe entre ellas y al tipo de bosque que condicionan la baja probabilidad de presencia de una especie en ambas parcelas.
Importancia ecológica (IVI’s)
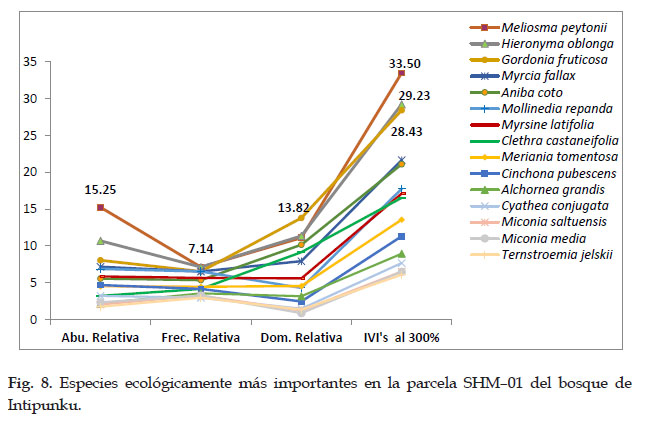
En la parcela SHM–01 del bosque de Intipunku, las especies ecológicamente más importantes son Meliosma peytonii con el 33,50 % de IVI’s, Hieronyma oblonga con el 29,23 % de IVI’s, Gordonia fruticosa con el 28,43 % de IVI’s, Myrcia fallax con el 21,67 %, Aniba coto vel sp. aff. con el 21,09 %. En cuanto a la dominancia, frecuencia y abundancia, las especies Meliosma peytonii fue la más abundante, la especie Gordonia fruticosa fue la más dominante y las especies Meliosma peytonii y Hieronyma oblonga se presentaron con mayor frecuencia.
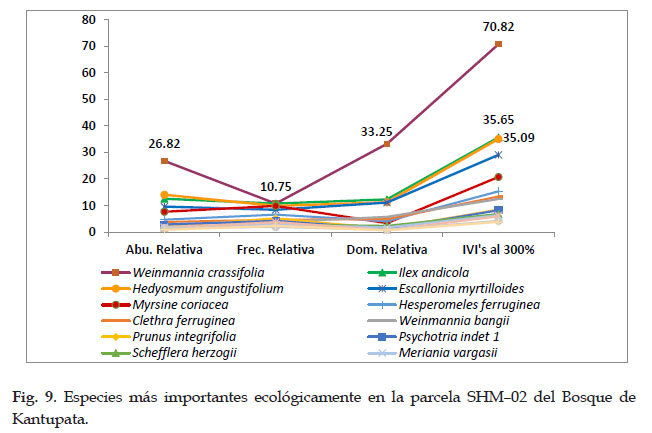
En la parcela SHM–02 de Kantupata se muestra a las 5 especies ecológicamente más importantes, de los cuales Weinmannia crassifolia con el 70,82 % de IVI’s fue la que resalta en importancia, seguida de Ilex andicola con 35,65 % de IVI’s, Hedyosmum angustifolium con 35,09 %, Escallonia myrtilloides con el 29,10 %, Myrsine coriacea con el 20,78 %, y el resto presento valores por debajo de estas especies.
En cuanto a la dominancia, frecuencia y abundancia, las especies Weinmannia crassifolia fue la más abundante y dominante, las especies Weinmannia crassifolia e Ilex andicola se presentaron con mayor frecuencia.
Discusión
Dinámica poblacional
El motor de la dinámica son las perturbaciones (mortalidad) que resultan en la formación de los claros, además de la regeneración (reclutamiento) y el crecimiento, que permiten mantener la estructura del bosque (Finegan, 1992). Estos procesos fueron analizados en el presente estudio, donde se determinó que las tasas de mortalidad fueron similares en ambas parcelas, pero las tasas de reclutamiento y renovación fue mayor en la parcela de Kantupata que se encuentra a mayor altitud, quiere decir que esta parcela presento mayor actividad poblacional. La diferencia de las tasas de reclutamiento entre las parcelas se puede deber a la gradiente altitudinal que existe entre ellas y al tipo de bosque, porque a mayor altitud (Kantupata a los 3200 m) el bosque es de dosel más bajo y de menor densidad poblacional, lo que permite mayor incidencia de la luz por presentar más claros y genera mayor competencia en el crecimiento, en Intipunku a los 2653m, ocurre lo contrario por lo se da un crecimiento lento de los árboles y está condicionado por los factores y condiciones ambientales del bosque, esto generó una baja tasa de reclutamiento.
Las tasas de mortalidad determinadas en ambas parcelas, corresponden a la mortalidad "Transfondo" que toma valores menores al 5 %, esto comprende eventos de mortalidad a pequeña escala como muertos en pie, rompimiento del tallo y volcamiento de raíz y ocasionan cambios graduales (Lugo & Scatena, 1996).
Para ser los primeros resultados de dinámica poblacional en los bosques estudiados, se muestra que ambas parcelas tienen un desequilibrio poblacional, al igual que en los bosques nublado de Venezuela (2200 – 2500 m) (Ramírez et al., 2012), esto se puede deber a que los bosques son jóvenes o bosques no maduros que están en proceso de crecimiento y recuperación como es el caso del bosque de Intipunku que en algún momento ha sufrido intervenciones antrópicas. Las tasas de reclutamiento y mortalidad del presente estudio son menores a lo reportado por Velásquez et al., 2012, en bosques andinos de Colombia, donde también se dio el desequilibrio entre dichas tasas. Los resultados de las tasas de mortalidad (1,39%) y reclutamiento (1,2%) reportados por Quinto et al., 2009, en un bosque pluvial tropical, son mayores a los resultados de Intipunku y menores a los de Kantupata, esta diferencia se debe principalmente al tipo de bosque donde se desarrolló cada investigación.
Biomasa aérea
Se presentó una clara diferencia de biomasa aérea entre ambas parcelas, e Intipunku tuvo mayor acumulación, esta variación se puede deber a la estructura, gradiente altitudinal y a la diferente composición florística que se presentó en cada una y al tipo de bosque donde se encuentra establecida cada una, ello concuerda con lo indicado por Meave et al.,
2006 y Álvarez, 2010, quienes mencionan que la variación de acumulación de C está relacionada con los cambios biofísicos registrados dentro del intervalo altitudinal, como su composición florística y estructural, y con lo mencionado por Homeier et al., 2010; López & Duque, 2010; Rangel-Ch et al., 2005, que en los bosques de montaña de los Andes (por encima de los 1800 msnm), la alta variación ambiental parece promover cambios abruptos en la estructura, composición y dinámica de la vegetación, incluso a escalas espaciales relativamente pequeñas.
Los valores de biomasa aérea reportados en el presente estudio, se encuentra dentro del rango de biomasa aérea reportado por Phillips et al., IDEAM 2011., en bosques montanos de Colombia, una variación de 105,3 Tha-1 a 295,1 Tha-1 entre los 1800 m y 3700 m de altitud, es similar a los reportado por Rodríguez et al. (2006), en el bosque de niebla en la reserva de la biosfera "El Cielo" en Tamaulipas, México y muy inferior al reporte de Velásquez et al. (2012), en bosques andinos de la Cordillera Occidental, Colombia.
Respecto a los estudios realizados en los bosques montanos del Perú, la biomasa estimada en la presente investigación es mayor a lo reportado por Girardin et al. (2010), en los altos Andes del Perú (Parque Nacional del Manu) entre los 2720 m a los 3025 m de altitud, donde registraron entre 47,03 TCha-1 y 65,90 TCha-1 de biomasa aérea. Lo registrado en Kantupata es similar a lo reportado por Gibbon et al. (2010) en bosques nublados y la puna de la Cordillera Oriental peruana (Parque Nacional del Manu), donde registró un promedio de 53,5 ± 5,7 MgCha-1 de carbono. Es menor a lo reportado por Cruzado (2010) en bosques altoandinos de la Concesión para Conservación Alto Huayabamba (CCAH), en el departamento de San Martín (3000–4600 m de altitud), quien registró 121,18 Tha-1 de biomasa aérea para el componente arbóreo, lo que se reporta para Intipunku es ligeramente superior a este valor. Mientras que, con lo reportado por Bellota, T. D. (2015), de 241,27 Tha-1 de biomasa aérea en el bosque montano (1391m de altitud) de la Reserva Comunal de "El Sira", el valor registrado en el presente estudio es muy inferior a este, esta diferencia básicamente se debe a la diferencia altitudinal, al modelo alométrico aplicado en la determinación de la biomasa aérea en cada investigación, al componente florístico, estructural (Meave et al., 2006; Álvarez, 2010) y a la relación de densidad especifica de madera con el DAP de las especies que componen cada bosque montano descrito anteriormente en las diferentes investigaciones mencionadas, por lo que regiones con altos valores de biomasa presentan mayor proporción de especies con alta densidad de madera (Baker et al., 2004).
Es necesario que haya más áreas de muestreo en los diferentes bosques montanos y tal vez que la estimación de biomasa aérea en cada uno se aplique con metodologías similares, para que haya menos sesgos y/o diferencias de estimación, además de la prioridad de evaluar los contenidos de carbono en la biomasa total (biomasa del suelo, de raíces, fuste, ramas y hojas) de los bosques montanos. En el presente estudio se presentó ciertas limitaciones, como solo 2 parcelas permanentes, para un área tan grande como del Santuario Histórico de Machupicchu, además estas parcelas reportan los primeros en el sector de Wiñaywayna.
Composición florística, IVI’s y diversidad
Se presentó una variación en la composición florística debido a la gradiente altitudinal de 635 m de separación entre las parcelas, al tipo de bosque montano y a las características en cada zona de vida, siendo la parcela de Intipunku de mayor número de individuos y especies que Kantupata. La diversidad alfa según los índices de Shannon y Simpson fue moderadamente alta en ambas parcelas, más la diversidad beta muestra una similitud muy baja entre ellas por compartir un mínimo de 7 especies. Esto demuestra que los bosques estudiados son muy diferentes y que a medida que incrementa la altitud, la riqueza de especies arbóreas disminuye.
Por la variabilidad de metodología aplicadas en los diferentes estudios, se compara con estudios realizados con la misma metodología aplicada en este estudio (arboles con DAP ≥ 10), los valores registrados en el presente estudio son menores a lo que reportaron Velásquez et al., (2012), en bosques andinos de la Cordillera Occidental, Colombia, donde registrarón 222 especies, 113 géneros y 60 familias en 1664 individuos en 2.0 ha de bosque, con 156 especies/ha, y es ligeramente menor a lo que registraron Ramírez et al. (2012), en bosque húmedo montano bajo (2200 y 2500 m de altitud) de la Cordillera de los Andes, Venezuela, donde registró 55 especies en 622 individuos en 1,5 ha, donde también registró a Myrcia fallax como la más representativa.
Respectos a estudios realizados en bosques montanos del territorio peruano, Palomino (1999), en el bosque nublado de San Pedro (Parque Nacional del Manú), reportó 192 especies, 91 géneros y 45 familias en 618 individuos en 1,0–ha, donde indica una alta diversidad y presencia de familias Moraceae (21,94%), Euphorbiaceae (17,7%), Melastomataceae (8,10%) como las más importantes, esto fue similar a lo registrado en Wiñaywayna del SHM, al igual que lo reportado por Rivera (2007) en la parte alta del Parque Nacional del Manu en la localidad de Wayquecha, donde registra a 2900 m de altitud una riqueza de 68 especies y 20 familias en 1 hectarea y donde las familias más diversas fueron Sabiaceae con 9 especies, Lauraceae (7) y Cyatheaceae (7), y las especies más representativas con mayor IVI fueron Weinmannia latifolia, Clusia cf. poepiggiana, Prunus integrifolia, Weinmannia crassifolia, Myrsine coriacea y Miconia livida; ambos lugares presentan similitud en la presencia de algunas familias y especies. Nina (2014), para los bosques nublados de Trocha Unión también del Parque Nacional del Manu reportó 43 especies, 22 géneros y 20 familias a los 3450 m y a 85 especies, 34 géneros y 26 familias a los 2750 m de altitud, estos valores fueron mayores a lo registrado en el presente estudio, pero comparten la presencia de algunos géneros y especies como Weinmannia, Cyathea, Myrsine, Miconia, entre otras. Estudios realizados próximos al área de estudio como el de Rojas & Peláez (2013), quienes mencionan una alta diversidad para la cuenca de Torontoy del Santuario Histórico de Machupicchu, estos valores son relativamente menores a lo reportado en el presente estudio, esta diferencia se debe a que el bosque de Intipunku ha sufrido extracción de muchas especies maderables en el proceso de construcción ferroviaria y se encuentran en proceso de recuperación, lo que mostro una menor diversidad. Otros estudios próximos, realizados con metodologías diferentes como los de Dueñas (1992), Monteagudo (1997), Galiano (2000), en los bosques de Wiñaywayna, mencionan un diversidad y composición florística muy similar a lo registrado para Wiñaywayna en el presente estudio.
En cambio, Galiano y Núñez (2011), quienes estimaron más de 550 especies de árboles distribuidas en 74 familias y 192 géneros para todo el Santuario Histórico de Machupicchu y zonas de amortiguamiento con Rubiaceae, Solanaceae, Asteraceae, Melastomataceae, Rosaceae, como las familias más representativas y los datos del presente estudio muestran un menor numero de especies, pues se desarrolló solo para los bosques de Wiñaywayna, más coinciden las familias y algunas especies reportadas en ambos estudios al igual que con los de Valenzuela et al. (2014), quienes reportaron más de 300 especies de árboles para el Santuario Histórico de Machupicchu y los describe en el libro "Arboles de Machupicchu Cusco–Perú". Hay una importante presencia de especies de helechos arbóreos dentro y fuera del SHM, esto es corroborado con los registros de Chávez (1994) quien registró dentro del Santuario Histórico de Machupicchu 4 especies y fuera de los límites del Santuario solo 3 especies, en el presente estudio registramos 7 especies de Cyatheas en el sector de Wiñaywayna.
Conclusiones
Las parcelas permanentes instaladas y monitoreadas en Wiñaywayna dentro del Santuario Histórico de Machupicchu, son las primeras para este Área Natural Protegida de gran importancia, en el presente estudio se logró determinar el primer registro de Biomasa aérea y dinámica poblacional al igual que la diversidad y composición florística a nivel de 1,0 ha de bosque, estos valores mostraron que los bosques montanos de Wiñaywayna son sumideros muy importantes de carbono al presentar altos valores de biomasa almacenada, los cuales pueden ser susceptibles a variaciones en el tiempo pues son datos iniciales, por lo que es necesario continuar con los monitoreos permanentes, además de promover más parcelas permanentes que abarquen mayor área de los bosques del SHM, lo que nos permitirán tener más datos y valores más certeros.
Agradecimientos
Al proyecto vía canon, "Diversidad florística y dinámica de los bosques de Wiñaywayna en el Santuario Histórico de Machupicchu", mediante el Vicerrectorado de Investigación de la Universidad Nacional de San Antonio Abad del Cusco. A RAINFOR y ForestPlots.net., al Jardín Botánico de Missouri, al convenio de Cooperación Científica MOBOT–UNSAAC, por brindar todas las facilidades necesarias para la ejecución de la presente investigación. Al Herbario Vargas (CUZ), a la Blga. Eufemia Machaca, residente de la estación Biológica de Wiñaywayna, por las facilidades y acceso a la Estación, al Sr. Macario Zuñiga por la asistencia brindada en la Estación Biológica de Wiñaywayna. A los colegas por su apoyo en campo: Miguel Pedraza Arando, Rosío Vega Quispe, Manuel Marca Zevallos, Jhon Gregory Quispe Almontes, Wilfredo Huamán Arqque, Elías Quispe, Marco Puértolas Yucra, Yanina Aymachoque, y Luz Danae Arqque Ch, todos ellos de la escuela profesional de Biología de la Universidad Nacional de San Antonio Abad del Cusco.
Contribución de autores
L. A., G. P; H. E. A. M. & W. Cha Levantarón la información de campo, sistematizarón, elaborarón la primera versión del artículo y aprobarón la publicación.
Conflictos de intereses
Los autores declaran no tener conflictos de intereses
Literatura citada
Álvarez, E.; A. Duque; J. Saldarriaga; K. Cabrera; G. de las Salas; I. del Valle; A. Lema; F. Moreno; S. Orrego & L. Rodríguez. 2012. Tree above-ground biomass allometries for carbon stocks estimation in the natural forests of Colombia. Forest Ecology and Management 267: 297-308. [ Links ]
Álvarez, A. G. 2010. Evaluación de la captura y estabilidad del carbon en un ecosistema de bosque mesófilo de montaña a lo largo del gradiente altitudinal en la Sierra Norte de Oaxaca. (Tesis de doctorado). Facultad de Ciencias. UNAM. 127 p. [ Links ]
Baev, P. & L. Penev. 1995. BIODIV: program for calculating biological diversity parameters, similarity, niche overlap, and cluster analysis. PENSOFT. Sofia-Moscú [ Links ].
Baker, T. R.; O. L. Phillips; Y. Malhi; S. Almeida; L. Arroyo; A. Di Fiore; T. Erwin; T. J. Killeen; S. G. Laurance; W. F. Laurance; S. L. Lewis; J. Lloyd; A. Monteagudo; D. A. Neill; S. Patino; N. C. A. Pitman; M. Silva & R. Vasquez. 2004. Variation in wood density determines spatial patterns in Amazonian forest biomass. Global Change Biology 10: 545-562. [ Links ]
Bellota, T. D. 2014. Diversidad florística, biomasa aérea y dinámica poblacional a través de la gradiente altitudinal en parcelas permanentes en la reserva comunal del Sira - Huánuco - Perú [ Links ].
Chave, J.; D. A. Coomes; S. Jansens; S. L. Lewis; N. G. Swenson & A. E. Zanne. 2009. Towards a worldwide wood economics spectrum. Ecology Letters 12: 351–366. [ Links ]
Chavez, H. W. 1994. Estudio Poblacional de Helechos Arborescentes en Wiñaywayna Santuario Histórico de Machupicchu y Cedropata. Seminario Curricular. Facultad de Ciencias Biológicas UNSAAC. [ Links ]
Cruzado, B. L. 2010. Determinación de las reservas de carbono en la biomasa aérea de los bosques altoandinos de la concesión para conservación alto Huayabamba – San Martín. [ Links ]
Cuesta, F.; M. Peralvo & N. Valarezo. 2009. Los bosques montanos de los Andes Tropicales. Una evaluación regional de su estado de conservación y de su vulnerabilidad a efectos del cambio climático. Quito. [ Links ]
Dueñas, L. H. 1992. Estructura diamétrica y diversidad arbórea en Wiñaywayna (Santuario Histórico de Machupicchu) Tesis para optar el Titulo de Biólogo, Facultad de Ciencias Biológicas UNSAAC. [ Links ]
Feinsinger, P. 2003. El diseño de estudios para la conservación de la Biodiversidad. Eds. FAN. Santa Cruz–Bolivia. 242p. [ Links ] Finegan, B. 1992. Bases ecológicas para la silvicultura. CATIE, Turrialba. 170 p. [ Links ]
Franco, L. J.; et al. 1989. Manual de Ecología, Edit. Trillas Segunda Edición México. PP, 14: 93-99 17: 114-119 18: 122-126 19:128-131 [ Links ]
Galiano, W. & P. Núñez. 2011. Sinopsis de la Especies Arbóreas del Santuario Histórico de Machupicchu y su Zona de Amor tiguamiento Revista El Antoniano N° 117, pág. 178. [ Links ]
Galiano, S. W. 2000. "Situación ecológico – ambiental del Santuario Histórico de Machupicchu". UNSAAC – Profonanpe, Cusco, Perú [ Links ].
Gentry, A. H. 1992. A Field Guide to the Families and Genera of Woody Plants of Northwest South America (Colombia, Ecuador, Perú) with supplementary notes on herbaceous taxa. 1ra ed. Washington, DC: Conservation International. 920 pp. [ Links ]
Gibbon, A.; M. Silman; Y. Malhi; J. Fisher; P. Meir; M. Zimmermann; G. Dargie; W. Farfán & K. Garcia. 2010. Ecosystem carbon storage across the grassland-Forest transition in the high Andes of Manu National Park, Peru. Ecosystems 13: 1097-1111. [ Links ]
Girardin, C. A. J.; Y. M. Malhi; L. E. O. C. Aragao; M. Mamani; H. W. Huaraca; L. Durand et al., 2010. Net primary productivity allocation and cycling of carbon along a tropical forest elevational transect in the Peruvian Andes. Global Change Biology. [ Links ]
Girardin, C. A. J.; W. Farfan-Rios; K. García; K. J. Feeley; P. M. Jørgensen; A. Araujo & Y. Malhi. 2013. Spatial patterns of above-ground structure, biomass and composition in a network of six Andean elevations transects. Plant Ecology & Diversity, 1-13. doi: 10.1111/j.1365-2486.2010.02235.x
Spracklen, D. V. & R. Righelato. 2013. Tropical montane forests are a larger than expected global carbon store. Biogeosciences Discuss, 10, 18893-18924. doi:10.5194/bgd-10-18893-2013 [ Links ]
Homeier, J.; S.W. Breckle; S. Gunter; R. T. Rollenbeck & C. Leuschner. 2010. Tree diversity, forest structure and productivity along altitudinal and topographical gradients in a species-rich Ecuadorian montane rain forest. Biotropica 42: 140-148. [ Links ]
Honorio, C. E. N. & T. R. Baker. 2010. Manual para el Monitoreo del ciclo del carbón en Bosques Amazónicos. Instituto de Investigaciones de la Amazonia Peruana/Universidad de Leeds. Lima, 54p. [ Links ]
Krebs, C. J. 1978. Ecology; the experimental analysis of distribution and abundance. Harper & Row (eds.). Nueva York. 678p.
López, W. & A. J. Duque. 2010. Patrones de diversidad alfa en tres fragmentos de bosques montanos en la región norte de los andes, Colombia. Rev. Biol. Trop. 58: 483-498. [ Links ]
Lugo, A. E. & F. N. Scatena. 1996. Background and catastrophic tree mortality in tropical moist, wet and rain forests. Biotropica 28:585-599. [ Links ]
Magurran, A. 1988. Ecological diversity and its measurements. Princeton University Press. New Jersey- U.S.A. 179 P. [ Links ]
Magurran, A. 1991. Diversidad ecológica y su medición. Vedra (Ed.). Barcelona – España. 54 – 70 p. [ Links ]
Magurran, A. 2004. Measuring biological diversity. Blackwell Science. Oxford – U.S.A. 256 P. [ Links ]
Meave, A. J.; A. Rincón & M. A. Romero-Romero. 2006. Oak forests of the hyper-humid region of La Chinantla, Northern Oaxaca range, México. In: Kappelle, M. (ed). Ecology and Conservation of Neotropical Montane Oak Forests. Ecological Studies 185. Springer-Verlag. Berlin, Heidelberg. pp:113-125. [ Links ]
Monteagudo, M. A. 1997. Evaluación de la diversidad arbórea en tres bosques representativos del Santuario Histórico de Machupicchu. Tesis para optar el título de Biólogo. Facultad de ciencias biológicas. Universidad Nacional de San Antonio Abad del Cusco – Cusco. [ Links ]
Moreno, E. C. 2001. "Métodos para medir biodiversidad" M&T – manuales y tesis SEA, vol1, Zaragoza, 84 pp.
Moreno, E. C.; I. Zuria; G. García; I. Sánchez; M. Castellanos; Martínez & A. Rojas. 2006. Trends in the measurement of alpha diversity in the last two decades. Interciencia 31: 67 – 71 p.
Nebel, G.; L. Kvist; J. Vanclay & H. Vidaurre. 2000. Dinámica de los bosques de la llanura alluvial inundable de la Amazonía Peruana: Efectos de las perturbaciones e implicancias para su manejo y conservación. Folia Amazónica Vol. 11, 65-97. [ Links ]
Nina, Q. A. 2014. Composición, Estructura y Regeneración Arbórea en dos Parcelas Permanentes del Bosque Nublado - Parque Nacional del Manu- Paucar tambo – Cusco. [ Links ]
Palomino, C. W. 1999. Diversidad y Asociación Arbórea del Bosque Nublado de San Pedro – Reserva de Biosfera del Manu. Tesis para optar el Titulo de Biólogo, Facultad de Ciencias Biológicas UNSAAC - Cusco - Perú [ Links ].
Phillips, J.; A. Duque; A. Yepes; K. Cabrera; M. García; D. Navarrete; E. Álvarez & D. Cárdenas. 2011. Estimación de las reservas actuales (2010) de carbono almacenadas en la biomasa aérea en bosques naturales de Colombia. Estratificación, alometría y métodos análiticos. pp. 68. IDEAM, Bogotá [ Links ].
Quinto, H. & R. Ibarguen. 2009. Mortalidad y Reclutamiento de Árboles en un Bosque Pluvial Tropical de Chocó. Rev. Facultad Nacional de Agronomía - Medellin., Vol. 62, num. 1, pp. 4855 - 4868. [ Links ]
Ramírez, A. H.; L. A. Torres & J. Serrano. 2002. Mortalidad y Reclutamiento de Árboles en un Bosque Nublado de la Cordillera de los Andes, Venezuela. ECOTROPICOS 15(2):177-184. [ Links ]
Rangel-Ch, J. O.; A. M. Cleef; S. Salamanca & C. Ariza. 2005. La vegetación de los bosques y selvas del Tatamá, p. 459-643. [ Links ]
In T. Van der Hammen, J. O.; Ch. Rangel & A. M. Cleef (eds.). La Cordillera Occidental colombiana – Transecto de Tatamá. Estudios de Ecosistemas Tropandinos, J. Cramer, Berlin-Stuttgart, Alemania. [ Links ]
Rivera, C. G. 2007. Composición florística y análisis de diversidad arbórea en un área de bosque montano en el Centro de Investigación Wayqecha, Kosñipata Cusco. Tesis de grado. UNALM- Lima. Pp. 121. [ Links ]
Rodríguez, L. R; P. Jiménez; C. Aguirre & E. Treviño. 2006. Estimación de carbono almacenado en un bosque de niebla en Tamaulipas, México. Ciencia- UANL. Monterrey, N. L. 9(2): 179-187. [ Links ]
Rojas, V. C. M. & T. Y. Peláez. 2013. Diversidad arbórea y cober tura vegetal en la cuenca de Torontoy, Santuario Histórico de Machupicchu – Cusco. [ Links ]
Sierra, C. A.; J. I. Del Valle; S. A. Orrego; F. H. Moreno; M. E. Harmon; M. Zapata; G. J. Colorado; M. A. Herrera; W. Lara; D. E. Restrepo; L. M. Berrouet; L. M. Loaiza & J. F.Benjumea. 2007. Total, carbon stocks in a tropical forest landscape of the Porce region, Colombia. Forest Ecology and Management, 243, 299-309. [ Links ]
Ureta, A. M. 2009. Diferencias Altitudinales de Contenido de Carbono y Biomasa Arbórea en el Parque Nacional Yanachaga Chemillén, Pasco-Perú [ Links ].
Valenzuela, G. L.; M. A. Monteagudo; H. G. Calatayud; V. J. Farfán; M. E. Suclli & Ch. I. Huamantupa. 2014. Arboles de Machupicchu Cusco – Perú. Jardín Botanico de Missouri a través de RAINFOR (Red Amazonica de Inventarios Forestales). [ Links ]
Velásquez, R. J.; N. Maniguaje & A. Duque. 2012. Diversidad y dinámica de un bosque subandino de altitud en la región nor te de los Andes colombianos. Rev. Biol. Trop. (Int. J. Trop. Biol. ISSN-0034-7744) Vol. 60 (2): 943-952. [ Links ]
Weberbauer, A. 1945 (1911). "El mundo vegetal de los Andes Peruanos". Ministerio de Agricultura, Lima. [ Links ]
Wolda, H. 1983. Diversity indices and tropical cockroaches. Oecologia 58: 290 – 298 p. [ Links ]
Zanne, A. E.; G. López-González; D. A. Coomes; J. Ilic; S. Jansen; S. L. Lewis; R. B. Miller; N. G. Swenson; M. C. Wiemann & J. Chave. 2009. Global Wood density database. Dryad. Identifier. [ Links ]
Recibido: 20-I-2018
Aceptado: 28-II-2018
Publicado online: 30-VII-2018
Publicado impreso: 30-VIII-2018

