










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkArnaldoa
versión impresa ISSN 1815-8242versión On-line ISSN 2413-3299
Arnaldoa vol.26 no.1 Trujillo ene./abr. 2019
http://dx.doi.org/10.22497/arnaldoa.261.26109
ARTÍCULOS ORIGINALES
Distribución espacial de la regeneración de Weinmannia lechleriana (Cunoniaceae) en un bosque montano del sector San Alberto, Parque Nacional Yanachaga-Chemillén (Perú)
Spatial distribution of the regeneration of Weinmannia lechleriana (Cunoniaceae) in a montane forest of the San Alberto Sector, Yanachaga-Chemillén National Park (Peru)
Robin Fernández-Hilario
Herbario Forestal MOL, Facultad de Ciencias Forestales, Universidad Nacional Agraria La Molina, Av. La Molina s/n, La Molina, Lima, PERÚ. rfernandez@lamolina.edu.pe
Resumen
Se estudió la regeneración natural de Weinmannia lechleriana Engl. (Cunoniaceae) en un bosque montano del sector San Alberto (Parque Nacional Yanachaga-Chemillén) en la selva central de Perú. Se evaluó la regeneración natural alrededor de diez árboles maduros, mediante el establecimiento de cuatro transectos de 20 metros de longitud. Se encontraron un total de 276 individuos alrededor de los diez árboles; el 57,2% correspondió a la categoría de brinzal, el 40,9% a latizal y solo el 1,8% a la categoría de fustal. El promedio de individuos de regeneración natural fue de 27 por árbol evaluado y la mayor densidad de regeneración fue encontrada entre los tres y seis metros alrededor del árbol, para luego ir disminuyendo al incrementar la distancia. Además, se halló una mayor abundancia total de individuos en los transectos oeste y en árboles evaluados en ladera, pero sería necesario evaluar un número mayor de árboles para llegar a una conclusión al respecto. Finalmente, podemos indicar que Weinmannia lechleriana aparentemente no presentaría problemas de regeneración en el área de estudio y se recomienda proseguir con los estudios de esta especie.
Palabras clave: Cunoniaceae, Pasco, estado de conservación, regeneración natural, brinzal.
Abstract
We studied the natural regeneration of Weinmannia lechleriana Engl. (Cunoniaceae) in a montane forest of the San Alberto Sector (Yanachaga-Chemillén National Park) in the central jungle of Peru. The natural regeneration was evaluated around ten mature trees, through the establishment of four transects of 20 meters in length. A total of 276 individuals were found around the ten trees, 57.2% corresponded to the category of seedlings, 40.9% of the saplings and only 1.8% to the category of poles. The average of natural regeneration individuals was 27 per evaluated tree, and the highest regeneration density was found between three and six meters around the tree, and then decreasing with increasing distance. Furthermore, a greater total abundance of individuals was found in the western transects and in trees evaluated in slope, but it would be necessary to evaluate a greater number of trees in order to reach a conclusion. Finally, we can indicate that Weinmannia lechleriana apparently would not present regeneration problems in the study area and it is recommended to continue with the studies of this species.
Keywords: Cunoniaceae, Pasco, conservation status, natural regeneration, seedling.
Introducción
Una de las mayores dificultades para el manejo de los bosques tropicales lo constituye el desconocimiento de las técnicas apropiadas en el campo de la silvicultura. Entre ellos se puede mencionar la casi inexistencia de tratamientos que involucren la regeneración natural, lo que finalmente solo terminan por generar grandes barreras que dificultan la formación de macizos forestales naturales heterogéneos, aptos a su respectiva ordenación y consecuente plan de manejo.
El éxito del manejo de un bosque tropical depende en gran parte de la existencia de suficiente regeneración natural que asegure la sostenibilidad del recurso a través del tiempo. Es por ello, que su cuantificación y análisis permite tener indicios del estado de conservación de las poblaciones, así como también la identificación de sus estrategias nos permiten establecer criterios para el manejo de especies intolerantes (heliófitas) o tolerantes a sombra (esciófitas) (Lamprecht, 1990; Donoso et al., 2014).
Las especies esciófitas son más susceptibles a los disturbios y deben ser las de mayor atención en planes de manejo y aprovechamiento (Felton et al., 2006). Por otra parte, las heliófitas son favorecidas para regenerarse ante la ocurrencia de disturbios y formación de claros; éstas son muy importantes en procesos de sucesión natural de bosques, debido a que promueven la regeneración de algunas especies esciófitas, aunque también podrían tornarse invasoras y suprimir la regeneración de otras especies arbóreas (Díaz & Armesto, 2007). Los planes de manejo deberían tener un enfoque integral para la conservación de la diversidad de flora según sus gremios ecológicos. Para esto se debería ordenar el bosque, planear medidas silviculturales y de domesticación de acuerdo a las necesidades ecológicas de las especies.
En el Perú, especies como Weinmannia crassifolia Ruiz & Pav. y Weinmannia lentiscifolia C. Presl se distribuyen en zonas de bosque montano nublado (1500-3500 msnm), y en bosques maduros, aunque también se las puede observar en los estadios de sucesión tardía (Reynel et al., 2016). Esto se puede explicar, en algunas especies, por su carácter de esciófitas "totales" (como en Weinmannia microphylla Ruiz & Pav.) que les permite poder germinar, crecer y permanecer toda su vida bajo sombra, no requiriendo de claros y generalmente ocupando el estrato medio del bosque (Ayma-Romay & PadillaBarroso, 2009), de esta manera pueden aprovechar la sombra que proyectan las especies heliófitas que se encuentran en el dosel o las esciófitas que ya están establecidas en el bosque para poder dar inicio a su ciclo vital.
Con respecto al establecimiento de las plántulas, Jaimes & Rivera (1991) mencionan para el caso de Weinmannia tomentosa L. f., que las semillas son poco longevas y no dormantes, mientras que el porcentaje de sobrevivencia en plántulas es bajo para Weinmannia pubescens (MorenoBetancur & Cuartas-Hernández, 2015). Si bien el establecimiento de la regeneración natural de especies de Weinmannia presenta una aparente dificultad durante sus primeras etapas en el bosque, se debe tener en cuenta que varias especies poseen importancia económica (Cordero & Boshier, 2003; Reynel et al., 2016) o ecológica (Bohórquez et al., 2011; MorenoBetancur & Cuartas-Hernández, 2015), lo que las convierte en un grupo promisorio dentro de la restauración ecológica y el manejo de los bosques tropicales. En este contexto, el objetivo del presente estudio fue determinar el radio de dispersión de la regeneración de Weinmannia lechleriana Engl. y cuantificar el número de individuos por categoría de regeneración, con la finalidad de obtener los primeros indicios sobre la ecología y estado de conservación de esta especie.
Material y métodos
Área de estudio
Este estudio fue realizado en el Sector San Alberto dentro del ámbito del Parque Nacional Yanachaga-Chemillen (PNYCH), en el departamento de Pasco (Perú). Esta zona pertenece a un bosque muy húmedo montano bajo (bmh-M); ubicado en el flanco Oeste de la Cordillera Yanachaga, entre los 2380-3000 metros de altitud, en las coordenadas geográficas S 10º32´32´´ W 75º 21´38´´ y S 10º33´10´´ W 75º21´04´´. En este bosque es frecuente la presencia de lluvias y neblinas constantes, por lo que también seria denominado como bosque nublado, además la precipitación promedio anual es de 2132 mm y la temperatura promedio anual está entre los 13 y 14 ºC. En la cordillera existen varios pasos importantes llamados comúnmente abras; uno de ellos es el Abra Esperanza, que comunica la quebrada San Alberto y la cuenca del rio Izcosacín hacia el flanco Este (Rojas et al., 2012).
Distribución espacial de la regeneración
El patrón de dispersión de plántulas, brinzales, latizales y fustales se determinó utilizando 10 individuos adultos y emergentes en pie de Weinmannia lechleriana, evaluadas durante el mes de febrero del 2012. Para evitar el traslape entre las áreas de muestreo se seleccionaron árboles que estuvieran distanciados, entre 40 y 100 metros aproximadamente. Desde cada árbol se realizaron transectos en los cuatro puntos cardinales, donde cada uno tuvo una longitud de 20 m y se subdividieron en cinco subparcelas: (a) dos subparcelas de 3 m de longitud por 2 m de ancho, (b) una subparcela de 4 m de largo por 2 m de ancho y (c) dos de 5 m de longitud por 4 m de ancho (ver Fig. 1). Se utilizó esta estrategia con el fin de reducir el sesgo de una intensidad de muestreo mayor a la orilla del árbol que en la región exterior del mismo e intensificar el muestreo en las áreas donde se espera que haya mayor incidencia de jóvenes. Se muestrearon 60 m2 por transecto, para un total de 240 m2 por árbol, lo que nos dio una área total de 0,24 ha de muestreo en el sector estudiado.
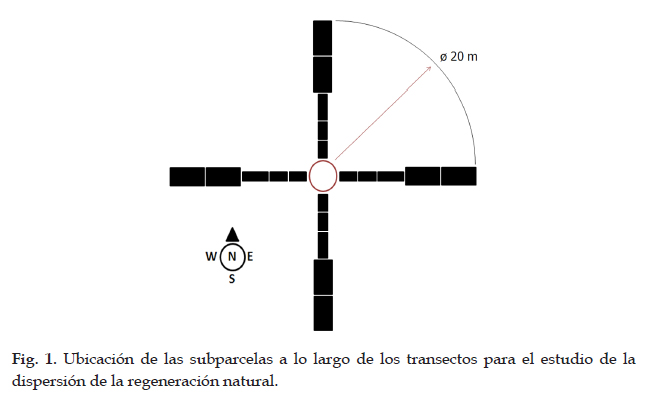
Categorías de regeneración
Dentro de cada subparcela se cuantificaron los individuos de regeneración natural presentes, según las siguientes categorías: (1) plántulas, todos aquellos individuos que se encontraron hasta la aparición de las primeras hojas verdaderas (eófilas), (2) brinzales, todas las plantas que estaban entre la aparición de las primeras hojas verdaderas y 1,5 m de altura, (3) latizales, individuos mayores de 1,5 m de altura y diámetros a la altura del pecho (DAP) hasta 9,9 cm, y (4) fustales, todos aquellos individuos con un DAP mayor a 9,9 cm.
Resultados
Los datos del radio de dispersión de las plántulas, brinzales, latizales y fustales obtenidos para Weinmannia lechleriana se resumen en el Cuadro 1. En total se encontraron 276 individuos de regeneración natural alrededor de los 10 árboles evaluados, dando una densidad de 0,115 ind/m2 (1150 ind/ha). El 57,2% de la regeneración natural perteneció a la categoría de brinzal, el 40,9% a latizal y solo el 1,8% a la categoría de fustal. Cabe señalar, que en el muestreo realizado no se registraron individuos en la categoría de plántulas. Además, se puede observar que el coeficiente de variabilidad (CV) es 0,41 y el promedio 27,6. Debido a que en los diez árboles muestreados la cantidad de regeneración natural puede oscilar entre 10 y 51 individuos. Mientras que para el caso de los brinzales, el coeficiente de variabilidad (CV) es 0,62 y el promedio 15,8; presentando una oscilación de individuos entre 5 y 27.
Los árboles evaluados presentaron diámetros a la altura del pecho (DAP) de 28 a 46 cm, y no se observó ninguna relación entre el DAP y la cantidad de individuos de regeneración natural.
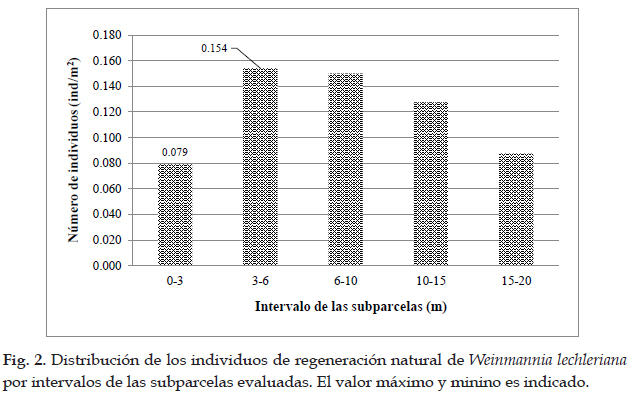
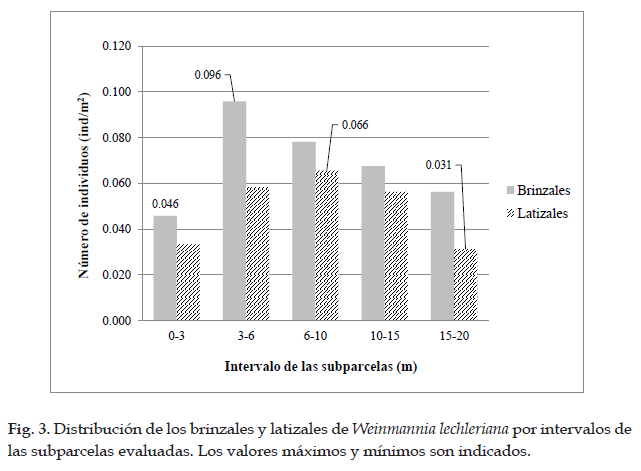
Encontrándose que el árbol WL03 (45 cm) registró la mayor regeneración (51 ind), mientras que el WL09 (39 cm) registró la menor regeneración (10 ind). Además, la mayor densidad de regeneración natural total (0,154 ind/m2) fue registrada en la subparcela comprendida en el intervalo de 3-6 metros, posterior a este rango los valores comienzan a disminuir gradualmente (Fig. 2). Para los valores de brinzales y latizales se observan tendencias similares, para el primero se presentó la mayor densidad (0.096 ind/m2) en el intervalo de 3-6 metros, mientras que para los latizales (0,066 ind/ m2) en el intervalo de 6-10 metros, para luego ir disminuyendo (Fig. 3).
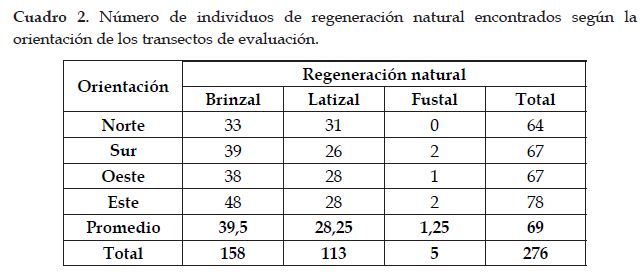
Con respecto a la orientación de los transectos de muestreo, se pudo apreciar que los valores obtenidos son similares, a excepción del transecto "Este" que presentó valores (78 ind) un poco mayores al resto (Cuadro 2). Además aquellos árboles evaluados que se encontraron ubicados en las laderas: WL02 (25 ind), WL03 (51 ind) y WL07 (40 ind) presentaron un promedio mayor de individuos de regeneración (38,7 ind) que el resto de árboles que se encontraban ubicados sobre las cimas ("cuchilla de aguas") (22,8 ind).
Discusión
La ausencia de plántulas durante la evaluación se debería a que Weinmannia lechleriana produce flores entre los meses de febrero y julio, como lo indican los especímenes botánicos procedentes de la zona de estudio depositados en el Herbario HOXA, y que la regeneración natural que se produjo el año anterior ya se encuentran en la categoría de brinzal o latizal. Se ha registrado que en otras especies de Weinmannia la etapa de floración pude ocurrir durante uno o siete meses (Palomino & Barra, 2003; Reynel et al., 2016), y que el proceso de maduración de frutos y diseminación de semillas se produce en los dos meses posteriores (Palomino & Barra, 2003). Además, la viabilidad de las semillas in situ para Weinmannia es corta (Jaimes & Rivera, 1991) y el porcentaje de germinación bajo (Josph & Delva, 2016), lo que limitaría la posibilidad de poder registrar plántulas durante un largo intervalo de tiempo. Asimismo, si bien el porcentaje de sobrevivencia de plántulas en viveros puede ser variable, registrándose aproximadamente hasta un 70% para Weinmannia lentiscifolia (Palomino et al., 1991) y un promedio inferior al 20% para Weinmannia pubescens Kunth (Moreno-Betancur & Cuartas-Hernández, 2014); en condiciones naturales Weinmannia poseería una alta mortandad de su regeneración debido probablemente a factores bióticos (competencia) o abióticos (humedad, luz), dando como resultado valores inferiores (1150 ind/ha) en comparación por ejemplo a los hallados para Weinmannia microphylla (aprox. 3000 ind/ha) en bosques poco intervenidos (Ayma-Romay & Padilla-Barroso, 2009).
Uno de los factores que pudo haber generado la baja densidad de regeneración naturalfuelagranproliferaciónde"pacales" en la zona de estudio (principalmente en los claros). Aunque la competencia entre especies diferentes es un factor negativo que puede llevar hacía la reducción de la supervivencia o crecimiento (Callaway, 2007), se pudo observar que existía una gran cantidad de individuos adultos de Weinmannia lechleriana bien distribuidos. Probablemente las categorías menores de regeneración (brinzales y latizales) sean tolerantes a sombra pero a un determinado tamaño requieran mayor cantidad y calidad de luz. Esto explicaría porque se registraron muy pocos individuos en la categoría de fustal, lo que también ha sido mencionado previamente (Palomino & Barra, 2003; Montes, 2011), al indicar que algunas especies de Weinmannia poseen una estrategia de crecimiento donde requiere semiluz para una buena germinación, y de sombra parcial en una primera etapa para luego convertirse en una especie heliófita.
Con respecto al radio de dispersión, a pesar de registrar una tendencia de la regeneración natural a disminuir mientras más se alejan del árbol evaluado no sería concluyente, porque las especies de Weinmannia posen semillas que pueden dispersarse fácilmente por acción del viento, debido a ser muy pequeñas y pubescentes (Rodríguez et al., 2006; Montenegro & Vargas, 2008). Esto concuerda con lo mencionado por Díaz & Vargas (2004), quienes reportan que Weinmannia tomentosa puede dispersarse grandes distancias, siendo la única especie dentro de su estudio que llego desde un bosque altoandino hacia pastizales abandonados. Podría ser que al aumentar la longitud de los transectos de evaluación se pueda seguir registrando regeneración natural, incluso en igual o mayor densidad.
Y sobre la cantidad de regeneración relacionada con la orientación de los transectos, solo para el caso de los brinzales se ve una mayor abundancia en el transecto Oeste (48 ind). Sin embargo, sería necesario muestrear más individuos con transectos más largos para obtener información que nos permita realizar alguna afirmación al respecto. En otras especies se han encontrado diferentes relaciones entre la influencia del sol en bosque liberados y la orientación de las fajas con regeneración natural (Rebottaro & Cabrelli, 2011; Canales-Springett et al., 2013). Mientras que confirmar el efecto de la topografía sobre la cantidad de regeneración natural también resultaría prematuro. Aunque, es bien cierto que en ambientes con déficit hídrico las laderas y su orientación generan diferentes condiciones microclimáticas que afectan la estructura de las poblaciones vegetales (del Castillo, 2000; López-Gómez et al., 2012; Revilla et al., 2015), resultados similares aún no han sido reportados en bosques montanos donde la humedad no es una variable limitante.
Conclusiones
La mayor densidad de individuos de regeneración natural de Weinmannia lechleriana se registró entre los 3-6 metros del árbol evaluado. Y a pesar de presentarse una mayor abundancia de individuos en los transectos Oeste y en árboles los evaluados en ladera, no es posible realizar una afirmación definitiva al respecto. Asimismo, podemos indicar que Weinmannia lechleriana aparentemente no presentaría problemas de regeneración en el sector de San Alberto, sin embargo, es probable que el establecimiento de fustales requiera mejores condiciones de luz. Finalmente, se recomienda realizar más estudios para analizar los posibles efectos de las variables ambientales (luz, viento, humedad) y la topografía sobre el establecimiento de la regeneración natural.
Agradecimientos
Al Jardín Botánico de Missouri – Oxapampa por darme la oportunidad de ejecutar este proyecto de investigación, a Rodolfo Vásquez, Luis Valenzuela y Rocío Rojas por el apoyo y asesoría durante la etapa de campo y las facilidades brindadas durante la estadía en las instalaciones del Herbario HOXA. Y a Ignacio Lombardi por sus comentarios en el borrador final del manuscrito.
Contribución de los autores
R.F.H.: Elaboró la metodología de evaluación, realizó el trabajo de campo y redactó el texto final.
Conflictos de intereses
El autor declara no tener conflictos de interés.
Literatura citada
Ayma-Romay, A. & E. Padilla-Barroso. 2009. Efecto de la tala de Podocarpus glomeratus (Podocarpaceae) sobre la estructura de un bosque de neblina en los Andes (Cochabamba, Bolivia). Rev. Peruana de Biología 16(1): 073-079. [ Links ]
Bohórquez, A. F.; D. Sanín & N. Walter. 2011. Estructura y composición arbórea de los bosques del Diablo (San Feliz, Salamina, Caldas), selva altoandina de la Cordillera Central Colombiana. Bol. Cient. Mus. Hist. Nat. 16(2): 39-52. [ Links ]
Callaway, R. 2007. Positive interactions and interdependence in plant communities. Springer. Dordrecht, Netherlands. 415 pp. [ Links ]
Canales-Springett, A. W.; A. Ceroni; G. Domínguez & A. Castillo. 2013. Respuesta de la regeneración natural de la Uncaria tomentosa (Willd) D.C. "uña de gato", al efecto de la luz en ecosistemas boscosos primarios intervenidos dentro del Bosque Nacional Alexander Von Humboldt, Pucallpa – Perú. Ecología Aplicada 12(2): 99-108. [ Links ]
Cordero, J. & D. H. Boshier (Eds.). 2003. Árboles de Centroamérica. Oxford Forestry Institute (OFI) & Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). 1079 pp. [ Links ]
del Castillo R. F. 2000. Composición y estructura de una nopalera bajo situaciones contrastantes de exposición de ladera y herbivoría. Boletín de la Sociedad Botánica de México 65: 5-22. [ Links ]
Díaz, M. & J. Armesto. 2007. Limitantes físicos y bióticos de la regeneración arbórea en matorrales sucesionales de la Isla Grande de Chiloé, Chile. Revista Chilena de Historia Natural 80: 13-26. [ Links ]
Díaz, R. & O. Vargas. 2004. Variación espacio temporal de la lluvia de semillas en pastizales abandonados de Alta montaña tropical (Reserva forestal municipal de Cogua). Acta Biológica Colombiana 9(2): 101-102. [ Links ]
Donoso, C.; M. González & A. Lara (Eds.). 2014. Ecología Forestal: bases para el manejo sustentable y conservación de los bosques nativos de Chile. Ediciones Universidad Austral de Chile. Valdivia, Chile. [ Links ]
Felton A.; A. M. Felton; J. Wood & D. B. Lindenmayer. 2006. Vegetation structure, phenology, and regeneration in the natural and anthropogenic tree-fall gaps of a reduced-impact logged subtropical Bolivian forest. Forest Ecology and Management 235: 186–193.
Jaimes, V. & D. Rivera. 1991. Banco de semillas y tendencias en la regeneración natural de un bosque altoandino en la regeneración de Monserrate (Cundinamarca, Colombia). Pérez-Arbelaezia 3(9): 3-35.
Joseph, A. & J. Delva. 2016. Respuesta germinativa de cuatro especies forestales nativas del Macizo del Cajas. Tesis para obtener el Título de Ingeniero Agrónomo. Facultad de Ciencias Agropecuarias, Universidad de Cuenca. Cuenca, Ecuador. 83 pp. [ Links ]
Lamprecht, H. 1990. Silvicultura de los trópicos: Los ecosistemas forestales en los bosques tropicales y sus especies arbóreas posibilidades y métodos para un aprovechamiento sostenido. Instituto de Silvicultura de Gottingen (GTZ). Antonio Carrillo (trad.). Eschborn, Alemania. 335 pp. [ Links ]
Lopéz-Gómez, V.; P. Zedillo-Avelleyra; S. Y. AnayaHong; E. González-Lozada & Cano-Santana, Z. 2012. Efecto de la orientación de la ladera sobre la estructura poblacional y ecomorfología de Neobuxbaumia tetetzo (Cactaceae). Botanical Sciences 90(4): 453-457.
Montenegro, A. & O. Vargas. 2008. Atributos vitales de especies leñosas en bordes de bosque altoandino de la Reserva Forestal de Cogua (Colombia). Rev. Biol. Trop. 56(2): 705-720.
Montes, C. 2011. Estado del conocimiento de Weinmannia tomentosa L.f. (encenillo) y algunas propuestas de estudio sobre su regeneración. Revista de Investigación Agraria y Ambiental 2(1): 45-53. [ Links ]
Moreno-Betancur, D. J. & S. E. Cuartas-Hernández. 2015. Sobrevivencia y crecimiento de plántulas de tres especies arbóreas en áreas de Bosque Montano Andino degradadas por ganadería en Colombia. Acta biol. Colomb. 20(2): 85-100. [ Links ]
Palomino, J. & M. Barra. 2003. Especies forestales nativas con potencial para reforestación en la provincia de Oxapampa y fichas técnicas de las especies de mayor prioridad. Pro Naturaleza, Programa Selva Central Oxapampa. Perú. 109 pp.
Palomino, J.; M. Barra; M. Bohórquez; G. Sosa & W. Hurtado. 1991. Ensayos silviculturales con especies y procedencias nativas en la Selva Central del Perú. Documento N°69. Instituto Nacional de Investigación Agraria – Deutsche Gesellschaft fur Technische Zusammenarbeit (GTZ). San Ramón, Perú. 58 pp.
Rebottaro, S. & D. Cabrelli. 2011. Regeneración natural de Pinus elliotti en claros silvícolas: dinámica poblacional durante siete años. Madera y Bosques 17(1): 49-70. [ Links ]
Reynel, C.; T. D. Pennington & R. T. Pennington. 2016. Árboles del Perú. The Natural Enviroment Research Council; Royal Botanic Garden Edinburgh; Darwin Initiative UK, Centro de Estudios en Dendrología – Fundación para el Desarrollo Agrario; Asociación Peruana para la Promoción del Desarrollo Sostenible. Lima, Perú. 1047 pp. [ Links ]
Revilla, I.; R. Fernandez-Hilario; S. Crespo & M. A. Astocaza. 2015. Diversidad y distribución de la familia Cactaceae y avifauna asociada en la Reserva Nacional de Lachay. Serie de Investigaciones CANDES 1: 10-35. [ Links ]
Rodríguez, M.; J. Puentes & F. Cortés. 2006. Caracterización temporal de la lluvia de semillas en un bosque nublado del cerro de Mamapacha (Boyacá – Colombia). Rev. Acad. Colomb. 30(117): 619-624.
Rojas, R.; M. Villalba; M. Vega; L. Valenzuela & R. Vásquez (Eds.). 2012. Jardín Botánico de Missouri: Curso Ecosistemas Andino-Amazónicos 2012. Programa For talecimiento Institucional y Capacitación. Oxapampa, Perú. 222 pp.
Recibido: 10-XII-2018
Aceptado: 20-II-2019
Publicado online: 10-IV-2019
Publicado impreso: 30-IV-2019

