










Servicios Personalizados
Revista

Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkArnaldoa
versión impresa ISSN 1815-8242versión On-line ISSN 2413-3299
Arnaldoa vol.26 no.1 Trujillo ene./abr. 2019
http://dx.doi.org/10.22497/arnaldoa.261.26112
ARTIGOS ORIGINAIS
Efeito do déficit hídrico e do aumento de temperatura sobre variáveis produtivas fisiológicas e bioquímicas do "cacau" Theobroma cacao L.
Effect of water stress and temperature increase on productive, physiological and biochemical variables of "cacao" Theobroma cacao L.
Ricardo Borjas Ventura1; Vanessa Mendoza Soto1; Alberto Julca Otiniano1 & Priscila Lupino Gratão2
1 Universidad Nacional Agraria La Molina, Departamento de Fitotecnia, Facultad de Agronomía, Lima, PERÚ. rborjas@lamolina.edu.pe
2 Universidade Estadual Paulista UNESP, Faculdade de Ciências Agrárias e Veterinárias, Depar tamento de Biologia Aplicada a Agropecuária, 14884-900 Jaboticabal, São Paulo, BRASIL.
Resumo
Tanto a seca quanto o aumento de temperatura serão incrementados pelas mudanças climáticas. Muitos cultivos serão afetados por esta drástica mudança no clima, entre eles está a cultura do "cacau" Theobroma cacao L. (Malvaceae). Tanto o déficit hídrico quanto o aumento de temperatura podem ocasionar danos fisiológicos e bioquímicos, que diminuem a produtividade desta espécie. Além disso, escassos trabalhos refletem o efeito destes estresses abióticos sobre o cacau. Portanto, torna-se necessário explorar o material genético que possa ser usado em condições de mudança climática. Neste sentido, esta revisão procura relacionar os trabalhos existentes e traçar estratégias sobre a importância desta cultura nos estudos que envolvam mudanças climáticas.
Palavras chaves: espécies reativas de oxigênio, seca, aquecimento global.
Abstract
Drought and temperature will be increased by climatic change. Many crops will be affected, for example, "cacao" Theobroma cacao L. (Malvaceae). Both water stress and temperature increase can cause physiological and biochemical damages, which diminish the productivity of this species. Moreover, there are few studies about the effect of these abiotic stresses on cacao. Therefore, it is necessary to explore the genetic material that can be used in conditions of climate change. In this sense, this review tries to relate the existing works and to draw strategies on the importance of this crop in the studies that involve climate change.
Key words: reactive oxygen species, drought, global warming
Introdução
O cacaueiro (Theobroma cacao L.) é uma das culturas mais importante do mundo, além, de ser cultivado em regiões onde os níveis de industrialização são baixos. Este cultivo abrange diversas regiões do mundo, entre elas América latina, sendo o Equador; o principal produtor.
O manejo desta cultura tem sido focada principalmente na sombra, na fertilização e no manejo de pragas, como a vassoura de bruxa (Moniliophthora perniciosa), "monilia" (Moniliophthora roreri) e um complexo de fungos do gênero Phytophthora. Porém, a cultura encontra-se exposta a diversos fenômenos climáticos como períodos de seca e altas temperaturas.
Pouca atenção se tem dado ao efeito destes agentes abióticos sobre Theobroma cacao. Segundo o IPCC, nos próximos anos teremos um aumento de temperatura de 2.3 °C entre 2046-2065 (Brown & Caldeira, 2017), que certamente estará acompanhado da seca.
Apesar do cacau ser plantado em regiões tropicais, muitas destas regiões têm períodos de baixa precipitação pluvial, cujo impacto na fisiologia, resposta antioxidante e produtividade não foram relatados. Estes estudos são extremamente importantes já que fornecem dados que podem ser usados em programas de melhoramento genético vegetal.
Muito trabalhos demostram que sob condições de altas temperaturas e déficit hídrico, a planta pode sofrer estresse oxidativo, que é causado por uma super produção de espécies reativas de oxigênio (ERO), tais como oxigênio singleto-1O , ión superóxido O˙¯, peroxido de hidrogênio- H2O2 e o radical hidroxila-•OH. (Mitller, 2017).
Cada ROS tem características diferentes, assim como os danos específicos que podem causar. O 1O , por exemplo, age sobre a membrana da célula, o O˙¯ ataca proteínas com o grupo Fe-S, enquanto o •OH, danifica membranas, DNA e proteínas (Barbosa et al., 2014). Neste ponto, é importante salientar que em condições normais ocorre a presença de ERO, mas estas são controladas eficientemente pelas plantas. Por outro lado, o efeito danoso da superprodução das ERO sobre diferentes compartimentos celulares é conhecido como estresse oxidativo.
Para lidar com a superprodução das ERO, e consequentemente evitar o dano oxidativo, a planta conta com amplo "arsenal" de mecanismos antioxidantes enzimáticos e não enzimáticos. Dentre os sistemas enzimáticos, podemos mencionar a Superóxide Dismutase (SOD, EC 1.15.1.1), Catalase (CAT. EC 1.11.1.6), Ascorbato Peroxidase (APX, EC 1.11.1.11), Glutationa Redutase (GR, EC 1.8.1.7), Monodehidroascorbato Redutase (MDHAR, EC 1.6.5.4) e Dehidroascorbato Reductase (DHAR, EC 1.8.5.1), enquanto o sistema de defesa não enzimático abrange compostos como Glutationa (GSH), Vitamina E, Ascorbato e carotenóides (Acosta et al., 2017; Asensi-Fabado & Munné-Bosch, 2010; Pintó-Marijuan & Munné-Bosch, 2013).
Deve-se salientar que o efeito dos estresses isolados, não induz a mesma resposta que a combinação de estresses. Apesar do aumento na acurácia das técnicas usadas para descrever o efeito do aumento de temperatura e do déficit hídrico sobre diversas espécies vegetais, estes estudos ainda são escassos em cacau. Porém, a sistematização dessa informação é importante para uma abordagem ampla do que existe na literatura e o que ainda carece de novos estudos, visando aumentar a produtividade desta cultura.
Produção de cacau em América Latina
O cacau é uma das culturas mais importantes do América Latina, dentre os principais países produtores estão o Equador e o Brasil. Com relação ao rendimento da cultura, a produção por hectare é baixa em alguns países, como por exemplo no Peru, onde reporta-se um rendimento de 800 kg ha-1 (AGRARIA, 2017), sendo 90 000 famílias diretamente relacionadas com seu plantio. (Selva y Sierra Exportadora, 2018). Outro grande produtor mundial de cacau é o Equador, com produção de 63% do cacau fino no mundo. Outros países importantes na produção de cacau na América do Sul são representados pela Colômbia, Venezuela e Brasil com rendimentos menores a 600 kg ha-1 (Tabela 1).
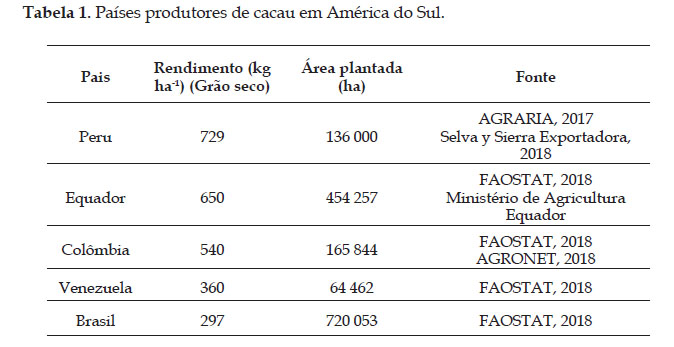
É importante considerar que a produção de cacau envolve a agricultura familiar, tendo o aumento de sua produtividade, forte impacto social. Como observado, a produtividade ainda é baixa, causada principalmente pelos efeitos climáticos da seca e do aumento da temperatura.
A frutificação do cacau ocorre durante todo o ano, o que favorece que este seja encontrado em uma mesma estação, em diferentes fases fenológicas. Deste modo, uma queda na precipitação pluvial pode ter impactos negativos na produção, não só por um estímulo na presença de pragas, mas também porque causar desequilíbrio fisiológico.
De acordo com diversas informações, as zona produtoras de cacau exibem períodos com queda na precipitação. Em algumas zonas produtoras de cacau no Peru, por exemplo, a maior precipitação concentra- se nos meses de novembro a março (MINAG, 2013). No caso de Colômbia, em geral, ocorre uma maior precipitação entre os meses de abril até novembro (Boletín Agroclimático, 2015). Enquanto que no Equador, a maior precipitação ocorre entre os meses de novembro a maio. Estes dados demostram a existência de períodos de baixa precipitação, os quais poderiam induzir o estresse oxidativo.
Efeito do estresse hídrico e térmico na resposta bioquímica do cacau
Antes de apresentarmos as respostas bioquímicas da planta de cacau, é importante situar ao leitor sobre o papel da resposta antioxidante perante situações de estresse, especificamente o déficit hídrico e ao aumento de temperatura.
Quando a planta encontra-se submetida a condições de estresse abiótico, como por exemplo o déficit hídrico e aumento de temperatura, ocorre a superprodução de espécies de oxigênio reativas (ERO), como oxigênio singleto (1O ), íon superóxido (O˙¯), peróxido de hidrogênio (H2O2) e o radical hidroxila (•OH). Cada ROS tem características diferentes, assim como os danos específicos que podem causar.
A Superóxido Dismutase (SOD) é considerada a primeira linha de defesa da planta. Esta enzima dismuta o ión superóxido (O˙¯) em peróxido de hidrogênio (H2O2) e água. Uma vez formado o H2O2, outras enzimas agem para transformá-la em água (como por exemplo, a CAT, APX e GPX). É importante ressaltar que se o H2O2 não é transformado em água, ele pode reagir com metais de transição (Fe ou Cu) e dar origem ao radical hidroxila (•OH), uma ERO extremamente danosa que danifica membrana plasmática, DNA, e proteínas (Sewelam et al., 2016), resultando no aumento da fluidez da membrana, mutações e perda de funcionalidade das proteínas.
Para atenuar o efeito negativo da superprodução de ERO, ocorre aumento da atividade das enzimas antioxidantes ou incremento na a síntese de compostos antioxidantes não enzimáticos. A resposta bioquímica de planta encontra-se dentre do que é conhecido como tolerância ao estresse. A tolerância envolve o ajuste osmótico e a síntese ou maior atividade de enzimas e moléculas antioxidantes.
Tanto o aumento de temperatura quanto a falta da água promovem uma maior atividade das enzimas antioxidantes, bem como, uma maior concentração de moléculas antioxidantes (Zandalinas et al., 2017), embora em situações extremas estas respostas não sejam suficientes para evitar possíveis danos celulares.
Existem poucos trabalhos sobre a resposta bioquímica da planta de cacau quando exposta ao déficit hídrico e ao aumento de temperatura. De acordo com Santos et al. (2014), em solos com potencial hídrico de -2 a -2,5 MPa as plantas de cacau apresentaram aumento na atividade das enzimas antioxidantes, Guaiacol Peroxidase e Polifenol Oxidase.
Efeito do estresse hídrico e térmico na produtividade do cacau
Poucos trabalhos demonstram o efeito da falta da água e do aumento da temperatura sobre o rendimento das plantas de cacau. A falta de atenção sobre o efeitos destes fenômenos climáticos pode ser estar relacionado ao seu local de plantio, nos trópicos, onde T. cacao não encontra condições de falta da água. Porém, a pouca informação existente sobre esse assunto mostra que sob condições de forte déficit hídrico, a produtividade da planta de cacau é afetada negativamente.
Tanto a seca quanto as altas temperaturas causam quedas no rendimento de vários cultivos como trigo (Zhang et al., 2013) e Vicia faba (Bishop et al., 2016). É importante destacar que o estresse hídrico não somente causa perdas no rendimento, como também causa uma queda na qualidade dos produtos (Alqudah et al., 2011). Uma das causas da perda do rendimento é a queda de flores. A queda das flores, causado pelo aumento de temperatura, é um fenômeno fisiológico complexo que envolve hormônios como o etileno (Djanaguiraman et al., 2011).
Segundo Hasanuzzaman et al. (2013), sob altas temperaturas, as plantas podem perder flores e frutos, e em condições extremas, podem até morrer. Além disso, este fenômeno pode ocorrer não somente com mudanças drásticas de temperatura, mas também com leves incrementos na temperatura. No caso do cacau, a seca pode provocar queda da florada, a qual traz uma menor quantidade de grão colhido. De fato, alguns trabalhos têm demonstrado que o déficit hídrico pode causar uma diminuição do rendimento em 45% da cultura (Schwendenmann et al., 2010), o qual converte a este tipo de estresse em uma ameaça importante para os produtores de cacau.
Mesmo assim, não só a queda de flores causa uma perda na produtividade, mas a excessiva temperatura também pode causas diminuição na massa seca dos grãos e do fruto. Daymond & Hadley (2008) relataram que acima de 22 °C, os grãos de cacau perdem peso.
Efeito do estresse hídrico e térmico na fisiologia do cacau
Um dos principais processos afetados tanto pelo aumento de temperatura quanto pelo estresse hídrico é a fotossíntese. A fotossíntese é um processo físico e bioquímico que fornece à planta de todos os insumos necessários para que possa se desenvolver.
Uma variável relacionada com a fotossíntese é a condutividade estomática. Segundo Daymond et al. (2011) a fotossíntese encontra-se fortemente correlacionada com a condutividade estomática. Dessa forma, ocorre intensa queda na condutividade estomática causada pelo déficit hídrico (Zhang et al., 2017). O fechamento estomático responde a uma adaptação da planta para não perder água. Entretanto, sob alta temperatura, as plantas aumentam a transpiração através da saída da água pelos estômatos.
Na planta de cacau "Criollo" sob déficit hídrico, a condutividade estomática (g) aumentou cerca de 2-3 vezes em períodos de chuva, comparado com a condutividade estomática quantificada em períodos de seca. Mesmo assim, a assimilação de CO2 e a transpiração também foram aumentadas com maior disponibilidade hídrica (Araque et al., 2012).
Existem diversas formas pelas quais o cacau consegue sobreviver a ambientes onde a disponibilidade da água é baixa. Uma dessas adaptações é o fechamento estomático. Segundo Carr & Lockwood (2011), solos com um potencial hídrico de -1.5 Mpa causaram o fechamento estomático em T. cacao. Resultados similares foram encontrados por García & Moreno (2015). Rada et al. (2005) notaram que em condições de falta da água a condutividade estomática foi reduzida, assim como a taxa de assimilação de CO2, com redução de 25%.
Embora ocorra o fechamento estomático a valores de -1.5 Mpa, o crescimento do cacau parece não estar comprometido com essa pouca quantidade da água no solo (Moser et al., 2010), o que sugere que a planta de cacau pode usar suas reservas para se manter estável durante a época de déficit hídrico.
O aumento da condutividade estomática em épocas de chuvas, e consequentemente, maiores taxas fotossintéticas, pode estar relacionada a maior produtividade favorecida nestes períodos.
Curiosamente, a eficiência fotossintética do fotossistema II (Fv/Fm) não foi modificada durante os períodos de baixa e alta precipitação pluvial (Araque et al., 2012). Isto poderia indicar uma adaptação dos clones estudados, para manter alta taxa fotossintética sob condições de pouca disponibilidade hídrica. Em outros estudos, Daymong & Hadley (2004), encontraram que cacaueiros em crescimento vegetativo demonstraram aumento na eficiência fotossintética em temperaturas de 23-25 °C, enquanto que em temperaturas superiores, a relação entre Fv/Fm, foi menor; o que poderia significar danos ao aparelho fotossintético.
Efeito do estresse hídrico e térmico no aspecto nutricional do cacau
Outro aspecto interessante é o efeito do déficit hídrico e o aumento de temperatura sobre o estado nutricional da planta de cacau. O estado nutricional da planta é muito importante já que os nutrientes podem atenuar o efeito de diferentes estresses sobre as culturas, como o Si e o Zn (Coskun et al., 2016; Candan et al., 2018).
Mesmo assim, tanto o déficit hídrico quanto o aumento da temperatura podem causar um desbalanço nutricional nas plantas, como de fato, foi observado em Brassica napus (Lee et al., 2016) e Zea mays (Zhao et al., 2015), sob condições de déficit, onde baixa absorção de S foi demonstrada.
Segundo Aranque et al. (2012), alterações significativas na absorção de N, P e K, no período seco e chuvoso ainda não foram encontradas. Porém, Santos et al. (2014) reportaram uma queda importante no teor de N, P, K, Ca e Mg no tecido vegetal de cacau, quando as plantas encontravam-se em solos com potenciais hídricos de -2 a -2,5 MPa. Tais perdas foram de 28, 22, 22, 69 e 56% comparados com a testemunha. Além disso, as concentrações de micronutrientes como Fe, Zn Cu e Mn também foram afetadas pela falta da água.
Efeito do estresse hídrico e térmico na resposta genética de cacau
As poliaminas são moléculas cuja síntese é aumentada quando a planta de cacau encontra-se perante de situações de estresse abiótico, especialmente sob seca e elevada temperatura. De acordo a literatura, podemos mencionar uma alta atividade dos genes TcODC, TcADC e TcSAMDC, os quais são altamente expressos quando a planta de cacau está em condições de déficit hídrico. Estes genes encontram-se ligados à síntese das poliaminas (Bae et al., 2008).
De acordo com Santos et al. (2014), os genes NCED5 e PP2C, sob déficit hídrico foram significativamente expressos pelo clone C.SUL 4. Os autores denotam que a super expressão de PP2C pode conferir alta sensibilidade ao C.SUL 4 em situações de déficit hídirco.
Perspectivas futuras
De acordo com a sistematização da informação do efeito da seca e do aumento de temperatura sobre a planta de cacau, os estudos presentes ainda são muito escassos, com foco principal nos aspectos fisiológicos, porém com pouca atenção a aspetos bioquímicos. Segundo a informação obtida, a planta de cacau mostra sensibilidade ao efeito da seca e do aumento de temperatura. Entretanto, salientamos a importância que novos estudos sejam realizados na cultura do cacau, frente às mudanças climáticas.
Agradecimentos
Os autores gostariam de agradecer o apoio da Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) e à Universidad Nacional Agraria La Molina (UNALM).
Contribuição dos autores
RBV, VMS, AJO e PLG participaram na ideia, a revisão crítica, a procura da informação bibliográfica e na correção da escrita deste trabalho.
Conflitos de interesse
Os autores declaram não ter conflito de interesse.
Referências
Acosta, J.; M. Ortuño; A. Bernal; P. Diaz; M. Sánchez & J. Hernández. 2017. Plant responses to salt stress: Adaptive mechanisms. Agronomy 7(18), 1-38.
Agele, S.; B. Famuwagun & A. Ogunleye. 2016. Effect of shade on microclimate, canopy characteristics and light integrals in dry season field-grown seedling. Journal of Hor ticultural Science 11(1), 47-56. [ Links ]
AGRARIA. 2017. El 93% de la producción peruana de cacao se concentra en 7 regiones. Disponible en http://agraria.pe/noticias/el-93-de-la-produccion- peruana-de-cacao-se-concentra-en-7-re-16171. [ Links ]
AGRONET. 2018. CACAO. Disponible en http://www.agronet.gov.co/Documents/Cacao.pdf. [ Links ]
Alqudah, A.; N. Samarah & R. Mullen. 2011. Drought stress effect on crop pollination, seed set, yield and quality. In: Lichtfouse, E. (Ed). Alternative farming systems, Biotechnology, drought stress and ecological fer tilisation. [ Links ]
Araque, O.; R. Jaimez; W. Tezara; I. Coronel; R. Urich & W. Espinoza. 2012. Comparative photosynthesis, water relations, growth and survival rates in juvenile criollo cacao cultivars (Theobroma cacao) during dry and wet seasons. Experimental Agriculture 48(4), 513-522. [ Links ]
Asensi-Fabado, M. & S. Munné-Bosch. 2010. Vitamins in plants: occurrence, biosynthesis and antioxidant function. Trends Plant Science 15, 582-592 [ Links ]
Barbosa, M.; M. Silva; L. Willadino; C. Ulisses & T. Camara. 2014. Geração e desintoxicação enzimática de espécies reativas de oxigênio em plantas. Ciência Rural 44(3), 453-460.
Bae, H.; S. Kim; M. Kim; R. Sicher; D. Lary; M. Strem; S. Natarajan & B. Bailey. 2008. The drought response of Theobroma cacao (cacao) and the regulation of genes involved in polyamine biosynthesis by drought and other stress. Plant Physiology and Biochemistry 46, 174-188.
Bae, H.; R. Sicher; M. Kim; S. Kim; M. Strem; R. Melnick & B. Bailey. 2009. The beneficial endophyte Trichoderma hamatum isolate DIS
219b promotes growth and delays the onset of the drought response in Theobroma cacao. Journal of Experimental Botany 60(11), 3279-3295.
Bishop, J.; S. Potts & H. Jones. 2016. Susceptibility of faba bean (Vicia faba L.) to heat stress during floral development and anthesis. Journal of Agronomy and Crop Science 202, 508-517. [ Links ]
Boletín Agroclimático. 2015. Contexto historico de las lluvias para el mes de Junio. Disponible en https://www.aclimatecolombia.org/download/notasprensa/Bolet%C3%ADnAgroclim%C3%A1tico6_Junio2015_Finalizado.pdf. [ Links ]
Brown, P. & K. Caldeira. 2017. Greater future global warming inferred from Ear th’s recent energy budget. Nature 552, 45-50.
Carr, M. & G. Lockwood. 2011. The water relations and irrigation requirements of cocoa (Theobroma cacao L.): A review. Experimental Agriculture 47(4), 653-676. [ Links ]
Candan, N.; I. Cakmak & L. Ozturk. 2018. Zinc- biofor tified seeds improved seedling growth under zinc deficiency and drought stress in durum wheat. Journal of Plant Nutrition and soil Science https://doi.org/10.1002/jpln.201800014. [ Links ]
Coskun, D.; D. Britto; W. Huynh & H. Kronzucker. 2016. The role of Silicon in higher plants under salinity and drought stress. Frontiers in Plant Science doi: 10.3389/fpls.2016.01072. [ Links ]
Daymong, A. & P. Hadley. 2004. The effects of temperature and light integral on early vegetative growth and chlorophyll fluorescence of four contrasting genotypes of cacao (Theobroma cacao). Annals of Applied Biology 145, 257-262. [ Links ]
Daynond, A. & P. Hadley. 2008. Differential effects of temperature on fruit development and bean quality of contrasting genotypes of cacao (Theobroma cacao). Annals of Applied Biology 153, 175-1785. [ Links ]
Daymond, A.; P. Tricker & O. Hadley. 2011. Genotypic variation in photosynthesis in cacao is correlated with stomatal conductance and leaf nitrogen. Biologia Plantarum 55(1), 99-104. [ Links ]
Djanaguiraman, M.; M. Prasad & K. Al-Khatib. 2011. Ethylene perception inhibitor 1-MCP decrease oxidative damage of leaves through enhanced antioxidant defense mechanisms in soybean plant grown under high temperature stress. Evironmental and Experimental Botany 17(2), 215-223. [ Links ]
FAOSTAT. 2018. Encontrado en http://www.fao.org/faostat/en/. [ Links ]
García, J. & L. Moreno. 2015. Respuestas fisiológicas de Theobroma cacao L. en etapa de vivero a la disponibilidad de agua en el suelo. Acta Agronómica 65(1), 44-50. [ Links ]
Hasanuzzaman, M.; K. Nahar; M. Alam; R. Roychowdhury & M. Fujita. 2013. Physiological, biochemical, and molescular mechanisms of heat stress tolerance in plants. International Journal of Molecular Sciences 14, 9643-9684. [ Links ]
Lee, B.; R. Zaman; J. Avice; A. Ourry & T. Kim. 2016. Sulfur use efficiency is a significant determinant of drought stress tolerance in relation to photosynthetic activity in Brassica napus cultivars. Frontiers in Plant Science. https://www.frontiersin.org/articles/10.3389/fpls.2016.00459/full.
Ministerio de Agricultura Ecuador. 2018. Rendimientos de cacao almendra seca (Theobroma cacao) en el Ecuador 2017. Disponible en http://sipa.agricultura.gob.ec/descargas/estudios/rendimientos/cacao/rendimiento_cacao_2017.pdf. [ Links ]
MINAG. 2013. Encontrado en http://infocafes.com/portal/wp-content/uploads/2016/03/cacao_completo.pdf. [ Links ]
Mitller, R. 2017. ROS are good. Trends Plant Science 22(1), 11-19. [ Links ]
Moser, G.; C. Leuschner; D. Hertel; D. Hölscher; M. Köhler; D. Leitner; B. Michalzik; E. Prihastanti; S. Tjitrosemito & L. Schwendenmann. 2010. Response of cocoa trees (Theobroma cacao) to a 13-month desiccation period in Sulawesi, Indonesia. Agroforestry Systems 79, 171-187. [ Links ]
Pintó-Marijuan, M. & S. Munné-Bosch. 2013. Ecophysiology of invasive plants: osmotic adjustment and antioxidants. Trends in Plant Science 18, 660-666. [ Links ]
Rada, F.; R. Jaimez; C. García-Núñez; A. Azócar & M. Ramírez. 2005. Relaciones hídricas e intrecambio de gases en Theobroma cacao var Guasare bajo períodos de déficit hídrico. Revista Facultad de Agronomía (LUZ) 22, 112-120. [ Links ]
Santos, I.; A. Almeida; D. Anhert; A. Conceição; C. Pirovani; J. Pires; R. Valle & V. Baligar. 2014. Molecular, physiological and biochemical response of Theobroma cacao L. genotypes on soil water deficit. PLOS ONE doi: 10.1371/journal. pone.0115746. [ Links ]
Selva y Sierra Exportadora. 2018. CACAO. Disponible en https://www.sierraexportadora.gob.pe/programas/cacao/que-significa.php. [ Links ]
Sewelam, N.; K. Kazan & P. Schenk. 2016. Global plant stress signalling: Reactive oxygen species at the cross road. Frontiers in Plant Science doi: 10.3389/fpls.2016.00187. [ Links ]
Schwendenmann, L.; E. Veldkamp; G. Moser; D. Hölscher; M. Köhler; Y. Clough; I. Anas; G. Djajakirana; S. Erasmi; D. Hertel; D. Leitner; C. Leuschner; B. Michalzik; P. Propastin; A. Tjoa; T. Tscharntke; O. Strateen. 2010. Effect of an experimental drought on the functioning of a cacao agroforestry system, Sulawesi, Indonesia. Global Change Biology 16, 1515-1530. [ Links ]
Zandalinas, S.; D. Balfagón; V. Arbona & A. Gómez-Cadenas. 2017. Modulation of antioxidant defense system is associated with combined drought and heat stress tolerance in citrus. Frontiers in Plant Science doi: 10.3389/fpls.2017.00953. [ Links ]
Zhang, X.; J. Cai; B. Wollenweber; F. Liu; T. Dai; W. Cao & D. Jiang. 2013. Multiple heat and drought event affect grain yield and accumulation of high molecular weight glutenin subunits and glutenin macropolymers in wheat. Journal of Cereal Science 57, 134-140. [ Links ]
Zhang, G.; M. Zhang; Z. Zhao; Y. Ren; Q. Li & W. Wang. 2017. Wheat TaPUB1 modulates plant drought stress resistance by improvement antioxidant capability. Scientific Reports doi: 10.1038/s41598-017-08181-w. [ Links ]
Zhao, R.; W. Guo; N. Bi; J. Guo; L. Wang; J. Zhao & J. Zhang. 2015. Arbuscular mycorrhizal fungi affect the growth, nutrient uptake and water status of maize (Zea mays L.) grown in two types of coal mine spoils under drought stress. Applied Soil Ecology 88, 41-49. [ Links ]
Recibido: 5-I-2019.
Aceptado: 25-II-2019.
Publicado online: 10-IV-2019.
Publicado impreso: 30-IV-2019.

