










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkArnaldoa
versión impresa ISSN 1815-8242versión On-line ISSN 2413-3299
Arnaldoa vol.26 no.2 Trujillo mayo/ago. 2019
http://dx.doi.org/10.22497/arnaldoa.262.26203
ARTÍCULOS ORIGINALES
Variabilidad morfológica de la "tara" Caesalpinia spinosa (Molina.) Kuntze (Fabaceae), en poblaciones naturales de Cajamarca: descriptores de fruto y semilla
Morphological variability of the "tara" Caesalpinia spinosa (Molina.) Kuntze (Fabaceae), in natural populations of Cajamarca: fruit and seed descriptors
Jim J. Villena Velásquez1, Juan F. Seminario Cunya2 y Miguel A. Valderrama Cabrera2
1 Escuela Profesional de Ingeniería Forestal Ambiental, Universidad Nacional Autónoma de Chota, PERÚ. jimjairo@hotmail.com
2 Programa de Raíces y Tubérculos Andinos, Universidad Nacional de Cajamarca, PERÚ. prtaunc@unc.edu.pe
Resumen
La "tara" o "taya" Caesalpinia spinosa (Molina) Kuntze (Fabaceae) es una especie sudamericana de importancia biológica, económica y cultural. Sus frutos y semillas son usados en medicina, alimentación y la industria, y provienen principalmente de poblaciones silvestres. El Perú es el principal exportador de "tara" y la región Cajamarca es el segundo productor en este país. Sin embargo, los estudios sobre variabilidad morfológica de la especie son escasos y confusos, lo cual limita su uso, estudio y conservación. Los estudios indican que los caracteres morfológicos discriminantes se encuentran en el fruto (vaina) y la semilla. Se reunieron 56 muestras de vainas de nueve provincias de la región Cajamarca. Mediante observación directa y con literatura especializada se establecieron once descriptores básicos de fruto y semilla, para determinar la variabilidad del germoplasma. Los datos se ingresaron a una matriz básica de datos y se procesaron con el programa NTSYS 2.2. Se identificaron siete morfotipos (clusters): Gigante, distribuida en Cajamarca, Contumazá y Chota; Blanca y Roja, distribuidas en las nueve provincias; Barbada, exclusiva de Celendín; Jancos, distribuida en Cajamarca, San Pablo y Santa Cruz; Ista, distribuida en Cajamarca y Contumazá; y Globosa, distribuida en Cajamarca. Los cinco primeros componentes principales explicaron 73 % de la varianza total y los caracteres que más aportaron a esta varianza fueron ubicación del hilo, forma de semilla, relación largo/ancho de vaina, color del lado opuesto al sol, apariencia superficial de la vaina, presencia de pelos glandulares en la vaina, peso de la semilla y relación largo/ancho de la semilla.
Palabras clave: Caesalpinia spinosa, "tara", variabilidad morfológica, descriptores, morfotipos, bosque natural.
Abstract
The "tara" or "taya" Caesalpinia spinosa (Molina) Kuntze (Fabaceae) is a South American species of biological, economic and cultural importance. Its fruits and seeds are used in medicine, food and industry, and mainly come from wild populations. Peru is the main exporter of "tara" and the Cajamarca Region is the second largest producer in this country. However, studies on the morphological variability of the species are scarce and confusing, which limits its use, study and conservation. Studies indicate that the discriminating morphological characters are found in the fruit (pod) and the seed. Fifty-six samples of pods from nine provinces of the Cajamarca region were gathered. Through direct observation and specialized literature, eleven basic descriptors of fruit and seed were established to determine the variability of germplasm. The data was entered into a basic data matrix and processed with the NTSYS 2.2 program. Seven morphotypes (clusters) were identified: Giant, distributed in Cajamarca, Contumazá and Chota; White and Red, distributed in the nine provinces; Bearded, exclusive of Celendín; Jancos, distributed in Cajamarca, San Pablo and Santa Cruz; Ista, distributed in Cajamarca and Contumazá; and Globose, distributed in Cajamarca. The first five main components explained 73 % of the total variance and the characters that contributed the most to this variance were yarn location, seed shape, pod length/width ratio, color of the opposite side to the sun, surface appearance of the pod, presence of glandular hairs in the pod, weight of the seed and long/wide relation of the seed.
Keywords: Caesalpinia spinosa, "tara", morphological variability, descriptors, morphotypes, natural forest.
Introducción
En el Perú existen 10 especies de Caesalpinia (Fabaceae), de las cuales, C. spinosa (Molina) Kuntze = Tara spinosa (Molina) Briton & Rose, conocida como "tara" o "taya" es económicamente, la más importante, cuya distinción fundamental de las otras nueve especies estaría en la presencia de acúleos o aguijones en el tallo, ramas y hojas; en el cáliz asimétrico y el sépalo abaxial más desarrollado y con dientes del ápice notorios (Dostert et al., 2009). Aunque se ha sugerido que la "tara" es una planta oriunda de Perú (Raimondi, 1857; Garro et al., 1997) no existen evidencias suficientes para sostener esta hipótesis. Además, la actualización de los endemismos de Perú indica que sólo C. chicamana Killip & Macbride y C. paipai Ruiz & Pav., son endémicas de este territorio (Baldeón et al., 2006).
La "tara" se distribuye en Venezuela, Colombia, Ecuador, Chile y Bolivia. También crece como planta introducida o subespontánea en Antillas, Cuba, Estados Unidos, Brasil, y Argentina (Ulibarri, 1996) y ha sido introducida al norte y este de África, la India e Islas Canarias (Brack, 1999; Dostert et al., 2009; Gagnon et al., 2016). En el Perú se adapta a la alta heterogeneidad de ambientes, desde la Costa hasta la Sierra, especialmente entre 1000 y 2600 msnm (Sagástegui, et al., 1996; Vásquez et al., 2010), donde forma bosques naturales muy antiguos; pero puede llegar a 3000 msnm (Rodríguez et al. 2017). Su estado de conservación en este país es vulnerable (DS O43-2006-AG).
Desde la antigüedad la planta sirve como leña, material de construcción y captadora de niebla. La vaina y la semilla se usan en medicina y colorantes y, son fuente de taninos, goma y germen para la in>dustria (Sagástegui et al., 1996; Brack, 1999; Cordero, 2015).
El Perú es el primer productor de vaina en Sudamérica. En 2015 y 2016 se reportaron ingresos por 31,4 millones y 32,5 millones de dólares, por exportaciones de 21,2 miles de toneladas y 21,4 miles de toneladas de vaina, respectivamente. En los mismos años, Cajamarca produjo 6,3 miles de toneladas y 2,7 miles de toneladas de vaina, respectivamente; ubicándose como la segunda región en producción, después de Ayacucho (SERFOR 2015 y 2016).
La "tara" que se comercializa proviene principalmente, de poblaciones silvestres y además, se ha mantenido en dos estados previos a la domesticación (Harlan, 1975): Como planta tolerada en las chacras, en donde provee sombra y sirve de referencia en la división y utilización de las mismas y, como planta fomentada, especialmente alrededor de las chacras, como parte del lindero y elemento de protección. Desde la primera década del 2000 en que se inicia la instalación de plantaciones con fines comerciales y como planta ornamental en las ciudades, se inicia también un proceso de domesticación acelerado, que probablemente, transformará la planta en sus características genéticas de fruto y semilla principalmente, puesto que son los órganos de mayor atención.
No se conocen los morfotipos y genotipos que contiene el germoplasma nacional y particularmente el de Cajamarca. El desconocimiento de la variabilidad fenotípica y genética, conlleva al inadecuado manejo y a la subutilización del recurso. Por ejemplo, se limita el conocimiento de las potencialidades, en relación al contenido de goma, taninos y otros compuestos de interés económico, para identificar y promover los materiales más productivos y delinear procedimientos de mejora genética. Al parecer, la variabilidad genética de la "tara" es estrecha (Balaguer et al., 2011), sin embargo, la planta muestra alta plasticidad fenotípica, esto explica su adaptación a múltiples calidades de sitio (Cordero, 2015). La identificación de morfotipos (y genotipos) facilitará el seguimiento del movimiento del germoplasma al interior del país, permitirá proponer estrategias de conservación y uso y, constituirá un elemento de respaldo contra la biopiratería (Singh et al., 2012; Singh et al., 2014).
Las descripciones botánicas antiguas y recientes de la "tara" resaltan que los caracteres diagnósticos más importantes se encuentran en la vaina (Ruíz & Pavón, 1777; León et al., 1994; Sagástegui et al., 1996; Ulibarri, 1996; Oliva et al., 2015; Sánchez et al., 2016; Gagnon et al., 2016) y la semilla (Ulibarri, 2008; Sagástegui et al., 1996; Bortoluzzi et al., 2007; Gagnon et al., 2016).
Del mismo modo, los primeros estudios sobre variabilidad morfológica de la "tara" comercializada, enfatizan el valor de la vaina y la semilla para este propósito. Así, Bonilla et al. (2016) encontraron seis semillas por fruto y la correlación peso de fruto y ancho de vaina fue buen estimador del peso de fruto. Orihuela (2014) usó seis caracteres cuantitativos de vaina y semilla y halló que la población de Ayacucho posee mejores características (que las de Cajamarca y Junín), porque sus frutos son más largos y pesados y las semillas son de mayor diámetro. Portal (2010), en Ayacucho, determinó cinco variedades de "tara": Almidón común, Almidón gigante, Morocho, Roja ayacuchana y Verde esmeralda; diferenciadas por la longitud de vaina, color de vaina y forma de semilla. Por su parte, Alemán et al. (2015) encontraron que los mejores caracteres discriminantes son de tipo cuantitativo de los mismos órganos y en 32 entradas bolivianas encontró tres grupos diferenciados.
En un estudio de Villar (2011) con "tara" de Cajamarca, determinó tres morfotipos: Roja o morocha, Blanca o Almidón y Celendina o Barbada. Los dos primeros, diferenciados por el color del fruto y el tercero por la presencia de pelos en la superficie de la vaina en estado verde.
Los antecedentes descritos son importantes, pero no hay claridad en la definición de los morfotipos, debido a la desuniformidad en los descriptores usados y porque en otros casos, se privilegian los caracteres cuantitativos, que son fuertemente influenciados por las condiciones ambientales, por lo que no siempre constituyen buenos descriptores. En este sentido, la pregunta ¿cuántos morfotipos de "tara" existen en cada región?, permanece sin respuesta.
El primer paso para el conocimiento del potencial genético de una especie es la caracterización morfológica, a través de descriptores estandarizados, fácilmente observables, con énfasis en aquellos que son altamente discriminantes, de alta heredabilidad y no afectados por las condiciones ambientales. El desarrollo de una lista básica de descriptores ayuda al registro sistemático y objetivo de los datos y facilita el intercambio de información entre investigadores y de éstos con los agricultores y otros usuarios (Hidalgo, 2003; Bioversity International, 2007).
Bajo estas consideraciones, las preguntas básicas que trató de responder la investigación fueron: (a) ¿cuáles son los descriptores morfológicos básicos del fruto y la semilla que permiten caracterizar el germoplasma de "tara" de nueve provincias de Cajamarca? y (b) ¿cuál es la variabilidad morfológica de "tara" de nueve provincias de Cajamarca?
Materiales y métodos
El estudio se realizó con vainas de "tara" colectadas en bosques y relictos de bosques de las provincias de Cajamarca, Cajabamba, Celendín, Contumazá, Chota, Hualgayoc, San Marcos, San Pablo y Santa Cruz, en donde se cosechan los mayores volúmenes para el mercado. En cada población identificada, con el apoyo de agricultores locales, se colectaron muestras de 50 a 100 frutos maduros en la planta. Cada árbol de donde se tomaron vainas fue georreferenciado y las muestras fueron numeradas consecutivamente, considerándose cada una como una entrada o accesión. El énfasis en las colectas estuvo dirigido a materiales reconocidos, por los lugareños como diferentes o probablemente diferentes, en cuanto a fruto y semilla, con atención a sus caracteres morfológicos.
En la determinación de descriptores se observó la población de vainas y se enfatizaron los caracteres morfológicos cualitativos, con el apoyo en la literatura especializada (Harrington & Durrel, 1981; Niembro, 1989; Ramírez & Goyes, 2004). En esta labor se siguieron los procedimientos recomendados por (Bioversity International, 2007). Cada descriptor consistió de (a) un nombre incluye objeto o ítem y una característica o atributo-, por ejemplo, color de vaina; (b) dos o más estados (valores que puede tomar cada descriptor); (c) un procedimiento o método de registro. Los colores fueron calificados con Colour Chart de RHS (The Royal Horticultural Society, 1995). También se tomaron en cuenta los pesos y la relación largo/ancho de vaina y semilla, porque en las pruebas previas mostraron valor discriminante. Esta inclusión se basó en que, algunas medidas de los órganos de las plantas mantienen relaciones alométricas estables, en cualquier condición o ambiente de crecimiento; de manera que, existe la seguridad de que la relación entre ellos no varía en el tiempo y en el espacio, aunque su valor absoluto puede variar (Gayon, 2000; Shinglenton, 2010).
Definidos los descriptores y sus estados, se aplicaron a cada entrada. Los datos fueron acopiados en una tabla de caracterización previamente probada. Los caracteres o descriptores altamente discriminatorios (según criterio de los investigadores) fueron marcados con un asterisco (*) (Bioversity International, 2007). El registro de los caracteres cuantitativos se hizo 50 vainas y 30 semillas por entrada e incluyeron: largo de vaina, ancho de vaina, espesor de vaina, peso de vaina con semillas, número de semillas por vaina, peso de vaina sin semilla, largo de semilla, ancho de semilla, espesor de semilla, peso de semilla por vaina, peso de semilla. Las dimensiones lineales se tomaron con pie de rey y los pesos se tomaron en balanza analítica con aproximación de tres dígitos.
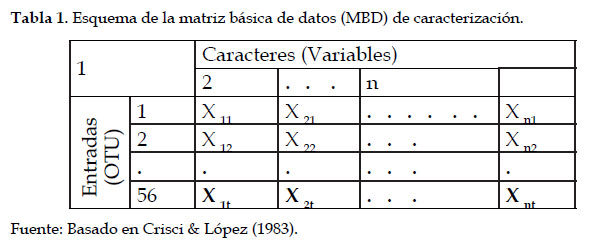
Los datos de los descriptores aplicados a cada entrada, fueron ingresados a una matriz básica de datos (MBD) elaborada en el programa Microsoft Excel versión 2010, la cual consistió de una tabla de doble entrada, en cuyas columnas se ubicaron los descriptores y en las filas las entradas con sus respectivos estados. De modo que, en cada casillero formado por la intersección de descriptor (columna) y entrada (fila) se ingresó el estado que correspondía a cada entrada para ese descriptor o carácter (Tabla 1). La información contenida en la MBD fue analizada con el programa NTSYS 2.2. (serie: OWIH4215J). Este análisis consistió de dos partes: el agrupamiento jerárquico de las entradas y el análisis de componentes principales (ACP). El primero permitió obtener un fenograma con los grupos o clusters que existen en el germoplasma en estudio. Este análisis permite establecer el grado de variabilidad de las entradas, o sea el número de morfotipos y, el nivel probable de duplicados, de acuerdo a su cercanía morfológica y de acuerdo al criterio de los investigadores. Cada morfotipo fue identificado con un nombre asignado, para facilitar la identificación y el manejo de la información. El ACP permitió conocer la relación entre los descriptores considerados y la semejanza entre las entradas (unidades taxonómicas operativas). Es decir, permitió conocer los descriptores que están asociados y que caracterizan en el mismo sentido o en sentido contrario y también, permitió conocer la distribución de los cultivares dentro del conjunto y los que más se parecen entre sí (Hidalgo, 2003).
El análisis de los datos cuantitativos por separado, siguió el mismo procedimiento que para los descriptores. La MBD se analizó mediante NTSYS 2.2 para obtener un fenograma que permitió agrupar a los materiales y luego se realizó el análisis de CP para conocer cuales, son los principales en la definición de la variación y qué caracteres son los que más aportan a estos CP. Además, se realizaron correlaciones entres los once caracteres cuantitativos observados, para conocer qué pares de caracteres mantienen relaciones significativas. En la interpretación se tomó en cuenta las asunciones de Martínez et al. (2009) que considera correlaciones regulares y perfectas cuando r ≥ 0.5 y r = 1, respectivamente; y de Rojas (2003) que considera que valores de r ≥ 0.4 corresponden a asociaciones que representan patrones naturales de variación.
Resultados y discusión
Lista de descriptores básicos para C. spinosa, basada en fruto y semilla
Los descriptores y sus estados quedaron definidos del modo como se expone a continuación y se proponen como una primera aproximación, para la caracterización de materiales cajamarquinos. La lista de descriptores aquí propuesta debe ser probada con otros materiales de Cajamarca y de otras regiones, con el propósito de configurar, en el mediano plazo, una lista de descriptores básicos, para C. spinosa en el Perú.
Lista de descriptores propuesta para C. spinosa, basada en fruto y semilla
1. Vaina (fruto)
1.1 Forma de vaina (FV)*
Registrada en vainas maduras y secas y, tomadas de la planta (no del suelo). Se toma en cuenta los bordes de ambos lados de la vaina.
1 Oblonga
2 Ligeramente falcada
3 Falcada

1.2. Apariencia superficial de la vaina (ASV)*
Registrada en vainas maduras y secas, y tomadas de la planta (no del suelo). Se observa la depresión entre las semillas, mejor en vista lateral de las vainas.
1 Aplanada.
3 Ligeramente globosa (en cada semilla)
5 Profundamente globosa (en cada semilla)

1.3. Pelos glandulares (PG) en la superficie de la vaina antes de la madurez*
Registrada en la vaina en estado verde, antes de la etapa de secado de maduración.
Se observa la superficie externa de las vainas.
0 Ausente
1 Presente

1.4. Color de vaina del lado hacia el sol
Registrada en vainas maduras y secas y, tomadas de la planta (no del suelo). Generalmente, es el lado de coloración más intensa. La toma del dato se hace con Colour Chart de The Royal Horticultural Society, 1995).
1 Rojo en ambos lados de la vaina (red 31 – 35)
3 Anaranjado rojizo en ambos lados de la vaina (Orange red 31 – 35)
5 Pajizo o crema o en ambos lados de la vaina (Yellow – Orange yelow 11 – 23)
1.5. Color de vaina del lado opuesto al sol
Registrada en vainas maduras y secas y, tomadas de la planta (no del suelo). Generalmente este lado es de coloración menos intensa. La toma del dato se hace con Colour Chart de The Royal Horticultural Society, 1995).
1 Rojo en ambos lados de la vaina (red 31 – 35)
3 Anaranjado rojizo en ambos lados de la vaina (Orange red 31 – 35)
5 Pajizo o crema, o en ambos lados de la vaina (Yellow – Orange yelow 11 – 23)
1.6. Relación largo/ancho de la vaina
Con medidas registradas en vainas maduras y secas y, tomadas de la planta (no del suelo), en una muestra de por lo menos 50 vainas, tomadas al azar. El largo se toma desde la base (punto de inserción al pedúnculo) hasta el ápice. El ancho se toma en el punto de mayor amplitud de la vaina.
1 Baja (< 4)
3 Intermedia (≥ 4 ≤ 6)
5 Alta (> 6)
2. Semilla
2.1. Forma predomínate de la semilla (FS)*
Registrada en semillas de vainas maduras y secas y, tomadas de la planta (no del suelo). En vista frontal y tomando en consideración el eje base-ápice
1 Obovada globosa
3 Obovada aplanada
5 Romboide

2.2. Color principal de la semilla
Registrada en semillas de vainas maduras y secas y, tomadas de la planta (no del suelo). La toma del dato se hace con Colour Chart de The Royal Horticultural Society, 1995).
1 Marrón (200A–B-C-D)
3 Marrón grisáceo (199A)
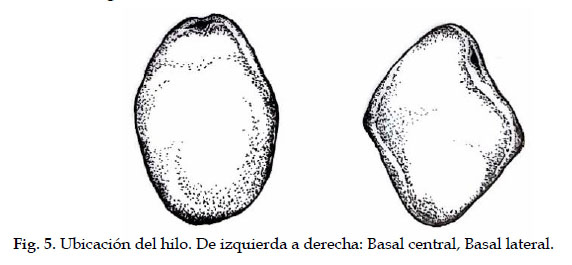
2.3. Ubicación del hilo*
Registrada en semillas de vainas maduras y secas y, tomadas de la planta (no del suelo)
1 Basal central
3 Basal lateral
2.4. Relación largo/ancho de la semilla
Con medidas registrada en semillas de vainas maduras y secas y, tomadas de la planta (no del suelo), en una muestra de por lo menos 30 semillas, tomadas al azar. El largo se toma desde el punto de inserción de la semilla a la vaina, hasta el ápice. El ancho se toma en posición transversal al largo y en el punto de mayor amplitud.
1 Baja (< 1)
3 Intermedia (≥ 1 ≤ 1.5)
5 Alta (> 1.5)
2.5. Peso promedio de la semilla (g)
Registrado en semillas de vainas maduras y secas y, tomadas de la planta (no del suelo). El peso promedio de cada semilla se obtiene de una muestra de por lo menos 30 semillas tomadas al azar.
1 Baja (< 1)
3 Intermedia (≥ 0.1 ≤ 0.2)
5 Alta (> 0.2)
Variabilidad morfológica de la tara, basada en el fruto y la semilla
El fenograma (Fig. 6), resultante del análisis de agrupamiento, indicó que a la distancia taxonómica de 0,00 nivel máximo de similitud, se forman 37 grupos, cuatro grupos conformados por dos entradas cada uno (E33, E41), (E15, E18), (E6, E12), (E32, E48), dos grupos conformados por tres entradas cada uno (E43, E34, E10) y (E14, E7, E3); un grupo conformado por cuatro entradas (E56, E50, E9 y E1) y el último conformado por siete entradas (E40, E45, E24, E17, E16, E13 y E11). A este nivel de análisis habría 38% de duplicados en el germoplasma en estudio.
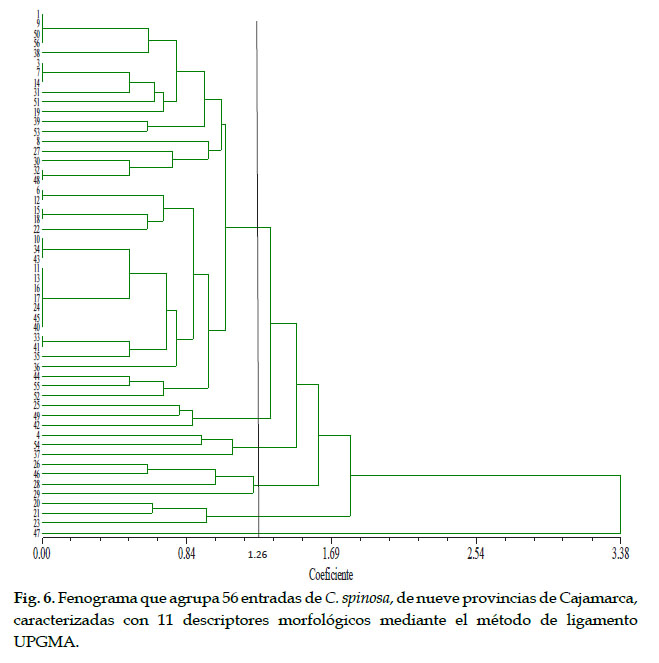
Tomando en consideración el coeficiente de similitud 1,26, se distinguen seis grupos. El sexto grupo contiene dos subgrupos visiblemente separados en el fenograma: el subgrupo (A) conformado por 22 entradas y el sub grupo B, conformado por 18 entradas. Estos dos subgrupos se diferencian en campo por el color de la vaina y son reconocidos por los agricultores de la región como "taya blanca" y "taya roja". Es decir, a este nivel de análisis y por razones de practicidad, en el germoplasma en estudio, de la región Cajamarca, tenemos siete morfotipos (88% de entradas duplicadas).
Morfotipos identificados en el germoplasma de tara de nueve provincias de Cajamarca
Una visión integral de los morfotipos de tara identificados en el germoplasma en estudio se presenta a través del nombre asignado, las entradas que los conforman y su distribución (Tabla 2). El nombre asignado corresponde a una de las características más relevantes o al sitio donde fue inicialmente encontrado el morfotipo. Las entradas conformantes corresponden a las entradas ubicadas en el mismo cluster dentro del fenograma (Fig. 5) y la distribución corresponde a las provincias de donde proceden las entradas que conforman cada morfotipo (Fig. 7).
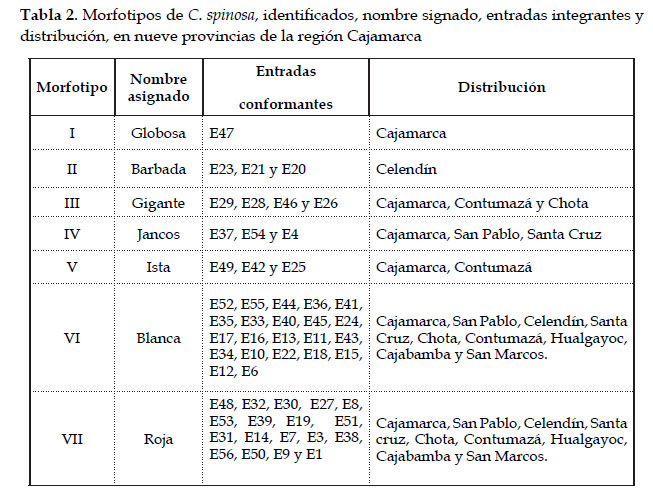
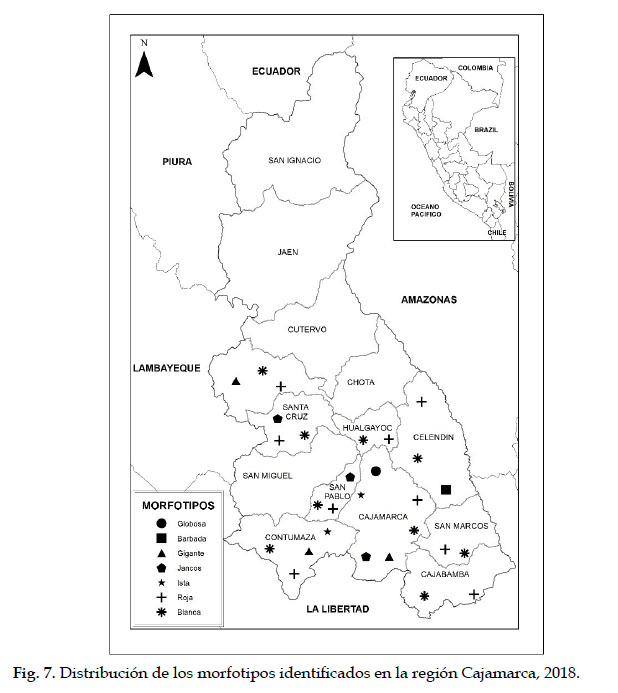
Los morfotipos identificados y su distribución en las provincias de colecta, permiten una primera aproximación a la variabilidad de la tara en la región. La distribución espacial de los morfotipos también es un indicio del estado de conservación en el ámbito de estudio: Así, los morfotipos Roja y Blanca se distribuyen en las nueve provincias, dos morfotipos (Globosa, Barbada) se distribuyen en una sola provincia, un morfotipo (Ista) en dos provincias, dos morfotipos (Gigante y Jancos) en tres provincias. En este sentido, es especialmente importante tomar en cuenta el morfotipo Globosa, del cual sólo se encontró un individuo en una sola localidad y Barbada, del cual se encontraron pocos individuos sólo en la provincia de Celendín. Este último sería un morfotipo endémico de Cajamarca, caracterizado por su vaina con pelos glandulares en la superficie, observables en estado verde.
Sin embargo, queda pendiente el estudio del germoplasma de cuatro provincias (Cutervo, San Miguel, Jaén y San Ignacio) (Fig. 8). Además, se deben profundizar las exploraciones en las provincias del sur, en donde se concentran las poblaciones más importantes de la especie. Por ejemplo, en Santa Cruz (Udima, Catache y Sexi), Chota (Lajas, Cochabamba, Huambos y Paccha), San Marcos (José Sabogal, Chancay, E. Villanueva e Ichocán), Celendín (Celendín y Utco), Cajabamba (Cachachi, Jocos y Cajabamba), Contumazá (Yonán, Tantarica, Guzmango, Chilete, Cupisnique y San Benito), San Pablo (Tumbadén, Jancos, Kuntur Wasi y San Bernardino). Asimismo, en los morfotipos identificados se deben realizar estudios sobre rendimiento de los componentes requeridos por el mercado (goma, taninos y vaina) y el análisis molecular para conocer la variación genética.
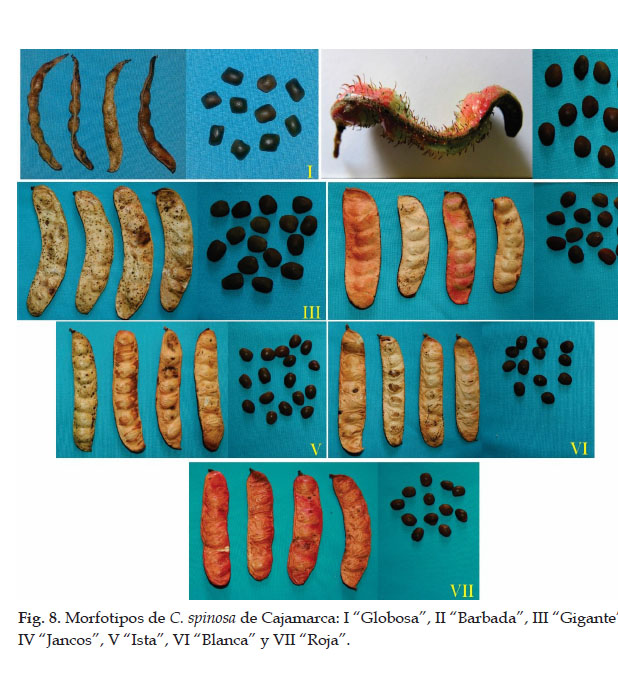
Las nuevas exploraciones deben permitir confirmar o ampliar el número de morfotipos en la región y una visión más clara sobre el estado de conservación de los mismos.
Análisis de componentes principales (CP), según caracteres morfológicos
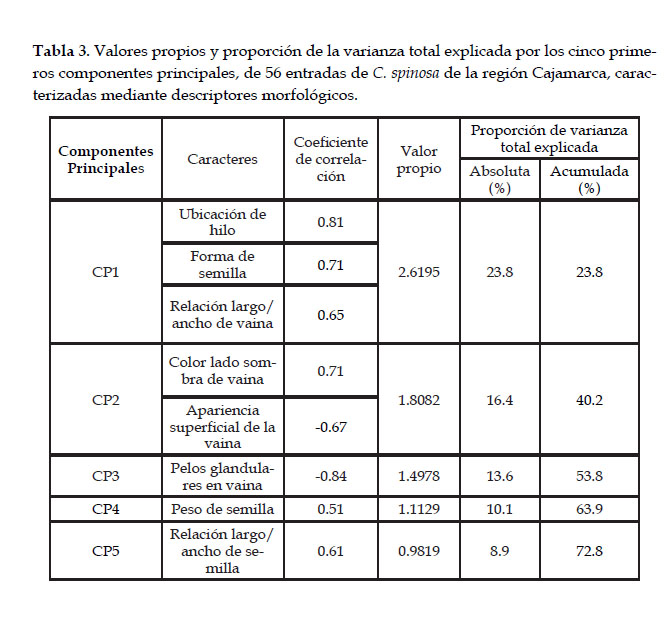
Siguiendo el criterio de Cliff citado por Rojas (2003) se seleccionaron los cinco primeros componentes principales que concentran el 73% de la variación total (Tabla 3). Las características asociadas a estos componentes son ocho, cuatro de semilla y cuatro de vaina. Es importante destacar que el primer componente principal explica el 23% de la variación y las características que contribuyen a esta fracción de la variación son la ubicación del hilo, la forma de la semilla y la relación largo/ancho de la vaina. Por otro lado, el cuarto componente explica el 10% de la variación y el peso de la semilla es el único contribuyente a esta variación. Para estudios posteriores se puede usar estas ocho características básicas para discriminar los materiales colectados.
Descripción de los morfotipos de tara de Cajamarca
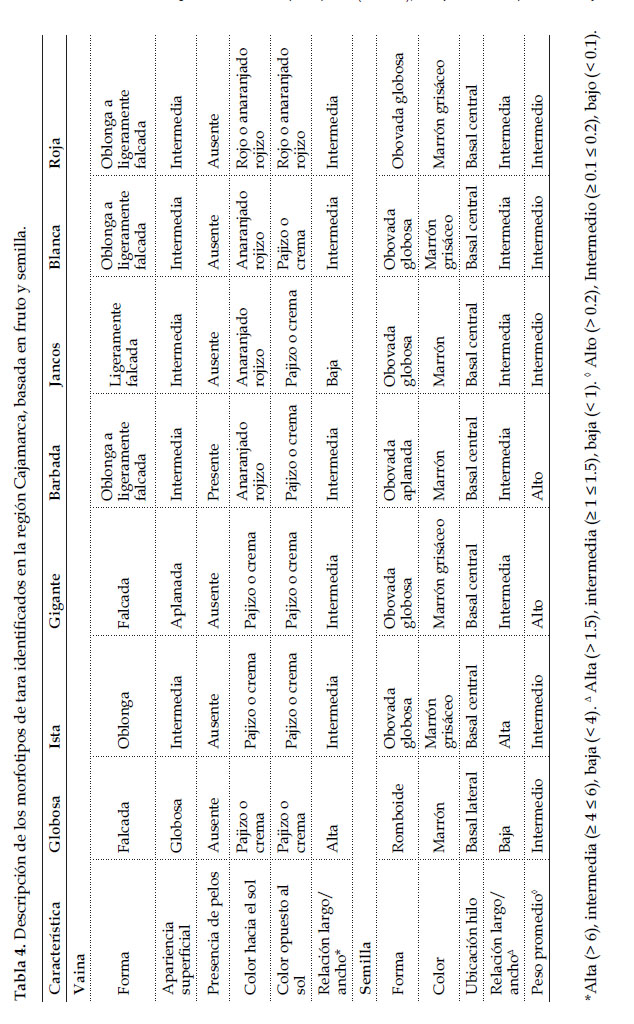
La descripción de los morfotipos identificados, basada en la caracterización con once descriptores morfológicos y en concordancia con los grupos derivados del agrupamiento se presenta a continuación (Tabla 4).
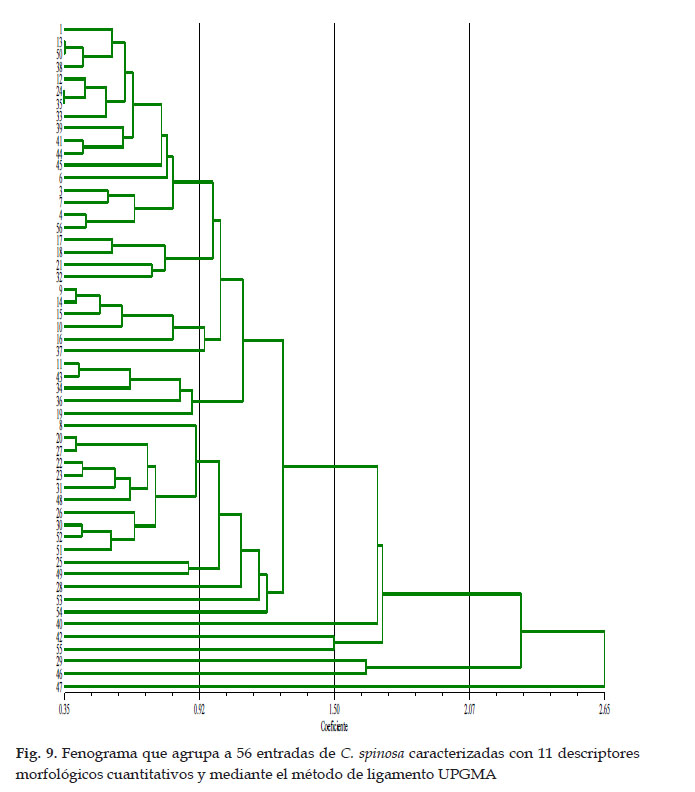
Agrupamiento de las entradas según los caracteres cuantitativos
A la distancia intermedia (coeficiente de similitud de 1,50) se diferencian siete grupos (Fig. 9), lo que indica que a este nivel de análisis el 88% de entradas son duplicadas. Seis grupos de una entrada cada una E47, E46, E29, E55, E42, E40. Un grupo conformado por cuatro sub grupos. (a) E54. (b) E53, E28, E49, E25, E51, E52, E30, E26, E48, E31, E23, E22, E27, E20, E8. (c) E19, E36, E34, E43, E11 y (d) E37, E16, E10, E15, E14, E9, E32, E21, E18, E17, E56, E4, E7, E3, E6, E45, E44, E41, E39, E33, E35, E24, E12, E38, E50, E13 y E1.
Análisis de componentes principales (CP), según los caracteres cuantitativos
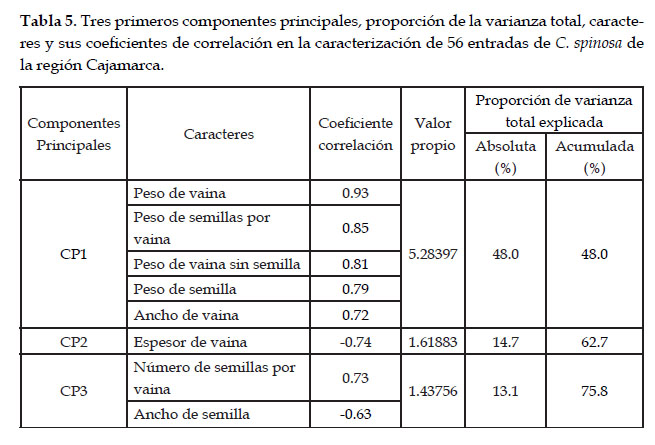
Los tres primeros componentes principales concentran el 76% de la variación total y los caracteres asociados a estos componentes son seis de vaina y dos de semilla. Destacan el peso de vaina (r = 0,93) y peso de semilla por vaina (r = 0,85) y en segundo lugar el peso de vaina sin semilla (r = 0,81) y el peso de la semilla (r = 0,79). Tres de estos caracteres también destacaron como descriptores en el estudio de Bonilla et al. (2016).
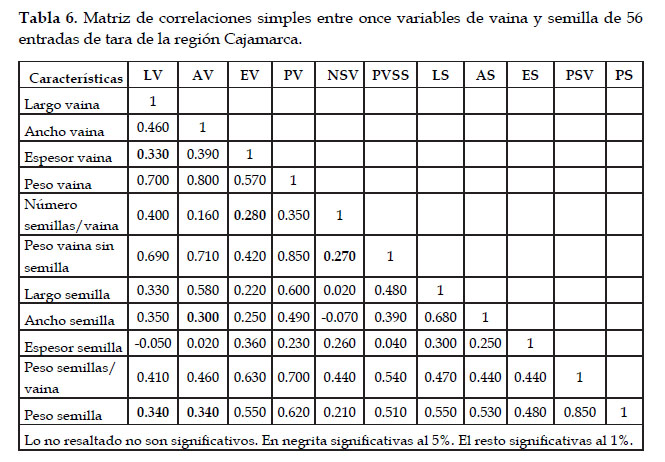
Correlaciones simples entre las dimensiones de vaina y semilla
La matriz de correlación entre once pares de características cuantitativas, siete referidas a la vaina y cuatro a la semilla (Tabla 6) se puede interpretar siguiendo a Rojas (2003) en su investigación sobre "quinua". Por un lado, observamos que 109 coeficientes fueron significativos (6 significativos a 5% y 103 significativos a 1%). Doce no fueron significativos. Por otro lado, el autor mencionado, considera que los coeficientes ≥ 0,40 corresponden a asociaciones que representan patrones naturales de variación. En este sentido, en primer lugar, es destacable la alta correlación de peso de vaina/ancho de vaina, peso de vaina sin semilla/peso de vaina y, peso de semilla/ peso de semillas por vaina (r ≥ 0,80). Otras correlaciones de alta significación (r ≥ 0,60 a r ≤ 0,71) son peso de vaina/largo semilla, espesor de vaina/peso semillas por vaina, peso de vaina/peso de semilla, largo vaina/peso de vaina, largo de vaina/peso de vaina sin semilla, peso de semillas por vaina/peso de vaina y, largo de semilla/ ancho de semilla. Asimismo, se confirma que las relaciones largo de vaina/ancho de vaina y largo de semilla/ancho de semilla, guardan relaciones alométricas significativas (r = 0,46 y r = 0,68, respectivamente) y constituyen buenos descriptores. Existen otras relaciones entre las medidas y pesos de vaina y semilla (peso semillas por vaina/peso vaina, peso de semilla/peso de semillas por vaina, peso de vaina sin semilla/peso de vaina, ancho de vaina/peso de vaina), que guardan relaciones altamente significativas y que podrían proponerse como buenos descriptores. Se sugiere seguir evaluando estas relaciones en nuevas colectas para confirmar la pertinencia de su inclusión como descriptores básicos.
Conclusiones
En 56 entradas de "tara" de nueve provincias de Cajamarca, se identificaron 11 descriptores de fruto (vaina) y semilla, básicos, para caracterizar el germoplasma.
Se determinaron siete morfotipos de "tara" en el germoplasma en estudio: Globosa (I), Barbada (II), Gigante (III), Jancos (IV), Ista (V), Blanca (VI) y Roja (VII).
Los caracteres más importantes para la discriminación de morfotipos son ubicación del hilo, forma de la semilla, relación largo/ancho de la vaina, color de vaina de lado opuesto al sol, apariencia superficial de la vaina, presencia de pelos glandulares en la vaina, peso de la semilla y relación largo/ancho de la semilla.
Los morfotipos Globosa se distribuye en Cajamarca; Ista se distribuye en Cajamarca y Contumazá; Gigante se distribuye en Cajamarca, Contumazá y Chota; Barbada es exclusiva de la provincia de Celendín; Jancos se distribuye en las provincias de Cajamarca, Santa Cruz y San Pablo y; Blanca y Roja se distribuyen en nueve provincias de la región Cajamarca.
Agradecimientos
A la Estación Experimental Baños del Inca, del Instituto Nacional de Innovación Agraria, por el apoyo logístico durante el desarrollo de la investigación. Al Ingeniero Humberto Valdez por el apoyo en el análisis estadístico. Al Ingeniero Ricardo Arribasplata por su orientación en la búsqueda de nuevos materiales. A los productores de "tara" de la región por su apoyo con información y material para el estudio.
Contribución de los autores
JJVV y JFSC, realizaron las colecciones, el trabajo de laboratorio y la redacción del informe.
MAVC, realizó los dibujos e hizo las revisiones del informe.
Conflicto de intereses
Los autores declaran no tener conflictos de interés
Literatura citada
Alemán, F.; C.Canelas & C. Ugarte. 2015. Validación de descriptores de Caesalpinia spinosa (Molina) Kuntze ("tara") en los valles interandinos de Bolivia. Revista de Agricultura 55:12-19. [ Links ]
Baldeón, S.; M. Flores & J. Roque. 2006. Fabaceae endémicas del Perú. En B. León, J. Roque, C. Ulloa, N. Pitman, P.M. Jørgesen y A. Cano (eds.). 2006. El libro rojo de las plantas endémicas del Perú. Rev. perú. biol. 13(2): 302-337.
Bonilla, H.; A. López; Y. Carbajal & M. Siles. 2016. Análisis de variables morfométricas de frutos de "tara" provenientes de Yauyos y Ayacucho para identificar caracteres agromorfológicos de interés. Scientia Agropecuaria 7 (3), 157 – 164. [ Links ]
Bioversity International. 2007. Guidelines for the development of crop descriptor lists. Bioversity Technical Bulletin Series. Bioversity International, Rome, Italy. 72 p. [ Links ]
Bortoluzzi, C.; S. Miotto; E. Biondo & T. Schiffino-Wittmann. 2007. Estudos morfológicos, citotaxonômicos e moleculares no grupo Caesalpinia L. sensu amplo: Caesalpinia, Hoffmanseggia e Pomaria no Sul da América do Sul. 47 p. [ Links ]
Brack, A. 1999. Diccionario enciclopédico de las plantas útiles del Perú. Programa de las naciones unidad para el desarrollo, Centros de estudios regionales andinos Bartolomé de Las Casas. pp. 88-89. [ Links ]
Cordero, I. 2015. Respuesta ecofisiológica de Caesalpinia spinosa (Mol.) Kuntze a condicionantes abióticos, bióticos y de manejo como referente para la restauración y conservación del bosque de nieblas de Atiquipa (Perú). Tesis doctoral. Facultad de Ciencias biológicas, Universidad Complutense de Madrid. 342 p. [ Links ]
Dostert, N.; J. Roque; G. Brokamp; A. Cano; M. I. La Torre & M. Weigend. 2009. Fctsheet: Datos botánicos de la "tara", Caesalpinia spinosa (Molina) Kuntze. Proyecto Perú Biodiverso, Desarrollo de monografía botánicas (Factsheets) para cinco cultivos peruanos. Lima, Perú. 9 p. [ Links ]
D.S. 043-2006-AG. 2006. Aprueban categorización de especies amenazadas de flora silvestre. El Peruano, Lima PE, jul. 13 (normas legales): 323527-323539. [ Links ]
Enríquez, G.; H. Cabanillas; C. Suárez & C. Salazar. 1997. Descripción y evaluación de los recursos genéticos. Técnicas para el manejo y uso de recursos genéticos vegetales. INIAP. Quito, Ecuador, 116-140. [ Links ]
Gayon, J. 2000. History of the concept of allometry. American Zoologist. 40(5): 748-758. [ Links ]
Gagnon, E.; A. Bruneau; C. E. Hughes; L. de Queiroz & G. P. Lewis. 2016. A new generic system for the pantropical Caesalpinia group (Leguminosae). PhytoKeys, 71:1-160. DOI: https://doi.org/10.3897/phytokeys.71.9203. [ Links ]
Garro, J. M.; B. Riedl & A. H. Conner. 1997. Analytical studies on tara tannins. Holzforschung 51(1997): 235-243. [ Links ]
Harlan, J. R. 1975. Crops and man. American Society of Agronomy, Crops Science Society of America. Madison, Wisconsin, US. pp. 63-64. [ Links ]
Harringtonm, H. D. & L. W. Durrel. 1981. How to identify plants. Ohio University. Swallow press, Chicago, US. 203 p. [ Links ]
Hidalgo, R. 2003. Variabilidad Genética y caracterización de especies vegetales. En T. Franco & R. Hidalgo. 2003. Análisis estadístico de datos de caracterización morfológica de recursos fitogenéticos. Instituto internacional de recurso fitogenéticos (IPGRI), Colombia. Boletín técnico N° 8:2-26. [ Links ]
León, B.; J. Gómez; J. Campos & R. Feux. 1994. Estudio taxonómico, ecológico, fitogenético y manejo agronómico de la "tara" Caesalpinia spinosa. Informe de investigación. Asociación Tecnológica y Desarrollo, Universidad Nacional Mayor de San Marcos. Lima Perú. 52 p. [ Links ]
Linares, J. R. 2014. Estudio de la diversidad genética de individuos de poblaciones silvestres de Caesalpinia spinosa (Molina) Kuntze "tara" mediante análisis de patrones electroforéticos de proteínas seminales. (tesis de pregrado). Universidad Nacional de San Maros. Lima. Perú. 64 p. [ Links ]
Martínez, R. M.; L. C. Tuya; M. Martínez; A. Pérez & M. Cánovas. 2009. El coeficiente de correlación de los rangos de Spearman. Caracterización. Rev haban cienc. Vol. VIII (2): 19. [ Links ]
Niembro, A. 1989. Semillas de plantas leñosas, morfología comparada. Editorial Limusa, S.A. de C.V. México, D.F. 50-51. [ Links ]
Orihuela, C. 2014. Evaluación de la diversidad genética de tres poblaciones de Caesalpinia spinosa procedentes de Cajamarca, Junín y Ayacucho mediante marcadores morfométricos de frutos y marcadores moleculares RAPD. (tesis de pregrado). Universidad Nacional Mayor de San Marcos. Lima. Perú. pp. 45 – 55. [ Links ]
Oliva, M.; R. Collazos; M. Chuquibala; I. Chuquizuta & C. Vogo. 2015. Caracterización morfológica de frutos y determinación del contenido de taninos en "tara" (Caesalpinia spinosa (Feuillée ex Molina) Kuntze) en las principales zonas productoras de la región Amazonas. Rev. Indes 1(2): 84-93. [ Links ]
Portal, E. 2010. Distribución altitudinal y elaboración de clave dicotómica y pictórica de biotipos de "tara" (Caesalpinia spinosa). Biológica Huamangensis 1(1), 1-5. [ Links ]
Raimondi, A. 1857. Elementos de botánica aplicada a la medicina y a la industria en los cuales se trata especialmente de las plantas del Perú. Segunda parte. Taxonomía, fitografía y geografía botánica. Tipografía Calle del Compas N° 202. Biblioteca Nacional de España. 222 p. [ Links ]
Ramírez, B. & A. Goyes. 2004. Botánica. Generalidades, morfología y anatomía de las plantas superiores. Popayán, Colombia: Universidad del Cauca.195 p. [ Links ]
Rojas, W. 2003. Análisis de la variabilidad genética en quinua. En T. Franco & R. Hidalgo. 2003. Análisis estadístico de datos de caracterización morfológica de recursos fitogenéticos. Instituto internacional de recurso fitogenéticos (IPGRI), Colombia. Boletín técnico N° 8. 89: 27-39. [ Links ]
Rodríguez, E. F.; S. Rodríguez; Y. Paredes & V. Rimarachín. 2017. Vegetales tintóreos más utilizados en la región La Libertad, Perú. Arnaldoa 24 (1): 331-332. [ Links ]
Ruiz, H. 1777. En la relación histórica del viage, que hizo a los reynos del Perú y Chile el botánico Hipólito Ruiz en el año de 1777 hasta el de 1788. Tomo I. [ Links ]
Sagástegui, A.; P. Lezama & E. Sánchez. 1996. Plantas promisorias: La "tara" o "taya". Arnaldoa 4(1), 57–65. [ Links ]
Sánchez, C.; E. Molinari-Novoa; E. Núñez-Linares & A. Arista. 2016. Advances on the floral morphology of Caesalpinia spinosa (Feuillée ex Molina) Kuntze "tara", a native tree to peruvian flora. The Biologist (Lima). Vol. 14 (1) 35-43. [ Links ]
SERFOR (Servicio Nacional Forestal y de Fauna Silvestre). 2016. Anuario forestal 2015. Ministerio de Agricultura. Lima, Perú. 218 p. [ Links ]
SERFOR (Servicio Nacional Forestal y de Fauna Silvestre). 2018. Anuario forestal y de fauna silvestre 2016. Ministerio de Agricultura. Lima, Perú. 107 p. [ Links ]
Shinglenton, A. 2010. Allometry: The study of biological scaling. Nature Education Knowledge 3(10), 2. [ Links ]
Singh, R. J.; A. Lobeda & A. O. Tucker. 2012. Medicinal plants—nature’s pharmacy. pp. 14–51. In: Singh, R. J. (ed.). Genetic Resources, Chromosome Engineering, and Crop Improvement. Medicinal Plants.
Singh, R. D., S. K. Mody; H. B. Patel; S. Devi; C. M. Modi & D. R. Kamani. 2014. Pharmaceutical biopiracy and protection of traditional knowledge. Int. J. Res. Dev. Pharm. L. Sci. 3(2): 866–871. [ Links ]
Ulibarri, E. A. 1996. Sinopsis de Caesalpinia y Hoffmannseggia (Leguminosae Caesalpinioideae) de Sudamérica. Darwiniana. 34(1-4): 299-348.
Ulibarri, E. A. 2008. Los géneros de Caesalpinioideae (Leguminosae) presentes en Sudamérica. Darwiniana. 46(1): 69-163. [ Links ]
Vásquez, L.; J. Escurra; R. Aguirre; G. Vásquez & L. Vásquez. 2010. Plantas medicinales del Norte del Perú. Fondo de Innovación Ciencia y Tecnología. Universidad Nacional Pedro Ruiz Gallo. Perú. Vol. 4. 345:384. [ Links ]
Villar, M. 2011. Identificación de morfotipos de taya en función a las características del fruto en la zona Sur de la región Cajamarca. Programa de Investigación en Sistemas Agroforestales Estación Experimental Agraria Baños del Inca. Proyecto CORECITI. Cajamarca. Perú. 25 p. [ Links ]
Weberbauer, A. 1945. El Mundo Vegetal de los Andes Peruanos. Ministerio de Agricultura, Lima. 77 p. [ Links ]
Recibido: 2-IV-2019
Aceptado: 15-VI-2019
Publicado online: 15-VIII-2019
Publicado impreso: 31-VIII-2019

