










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkArnaldoa
versión impresa ISSN 1815-8242versión On-line ISSN 2413-3299
Arnaldoa vol.26 no.2 Trujillo mayo/ago. 2019
http://dx.doi.org/10.22497/arnaldoa.261.26207
ARTÍCULOS ORIGNALES
Diversidad de especies de Porphyra y Pyropia (Bangiaceae, Rhodophyta) de Marcona (Ica, Perú) bajo la evidencia molecular
Species diversity in Porphyra and Pyropia (Bangiaceae, Rhodophyta) from Marcona (Ica, Peru) according to molecular evidences
Diego Márquez Corigliano1, Natalia Arakaki2, Patricia Gil Kodaka3, Florence Tellier4
1 Facultad de Ciencias, Universidad Nacional Agraria La Molina. Av. La Molina S/N, Lima, PERÚDepartamento de Ecología, Facultad de Ciencias, Universidad Católica de la Santísima Concepción. Alonso de Ribera 2850, Concepción, CHILE dimarquezcori@gmail.com
2 Banco de Germoplasma de Organismos Acuáticos, Instituto del Mar del Perú. Esquina Gamarra y General Valle s/n, Callao, PERÚ. narakaki@imarpe.gob.pe
3 Facultad de Pesquería, Universidad Nacional Agraria La Molina. Av. La Molina S/N, Lima, PERÚ pgilkodaka@lamolina.edu.pe
4 Departamento de Ecología, Facultad de Ciencias, Universidad Católica de la Santísima Concepción. Alonso de Ribera 2850, Concepción, CHILE ftellier@gmail.com
Resumen
La sistemática de Bangiales foliosas ha tenido cambios significativos a nivel global, debido a la inclusión de nuevos géneros, como Pyropia, y la incorporación de la evidencia molecular. En Chile, la aplicación de herramientas moleculares ha evidenciado una alta diversidad de especies de los géneros Porphyra y Pyropia, con delimitaciones de especies que no corresponden a las especies definidas con base en caracteres morfológicos. En el Perú, los registros históricos reconocen cinco especies de Porphyra; sin embargo, su estatus taxonómico aún no ha sido contrastado profundamente con la evidencia molecular. Por ello, este trabajo tuvo como objetivo explorar la diversidad local de las especies de los géneros Porphyra y Pyropia en Marcona, Perú, caracterizando 11 especímenes con los marcadores mitocondrial COI y cloroplastidial rbcL. El análisis filogenético identificó una especie de Porphyra (Porphyra sp. CHF) y tres especies de Pyropia (Pyropia orbicularis, Py. suborbiculata y Pyropia sp. CHI). No hubo coincidencia con los registros históricos de Porphyra. Tres de las cuatro especies encontradas tienen un rango de distribución que comprende Chile y Perú. Este trabajo es un primer avance hacia una caracterización de la diversidad específica del recurso algal conocido en Perú como "cochayuyo".
Palabras clave: COI, filogenia, identificación de especies, rbcL.
Abstract
The systematic of bladed Bangiales is suffering significant changes at a global level, due to the inclusion of new genera, such as Pyropia, and the incorporation of molecular evidence. In Chile, the application of molecular tools has shown a high diversity of species in genera Porphyra and Pyropia, with species delimitations that do not correspond to species defined on the basis of morphological characters. In Peru, historical records recognized five species of Porphyra, however, their taxonomic status has not yet been deeply contrasted with molecular evidence. Therefore, this work aimed to explore the local diversity of species of Porphyra and Pyropia in Marcona, Peru, characterizing 11 specimens with the mitochondrial and chloroplast markers COI and rbcL respectively. The phylogenetic analysis identified one species of Porphyra (Porphyra sp. CHF) and three Pyropia species (Pyropia orbicularis, Py. suborbiculata and Pyropia sp. CHI). There were no coincidences with the historical records of Porphyra. Three of the four species found have a distribution range encompassing Chile and Peru. This work is a first step towards a characterization of the specific diversity of the algal resource known as "cochayuyo" in Peru.
Keywords: COI, phylogeny, species identification, rbcL.
Introducción
A través de los análisis moleculares se han establecido nuevos géneros de algas rojas filamentosas y foliosas de la familia Bangiaceae, siendo muchos de ellos monotípicos (Sutherland et al., 2011; Sánchez et al., 2014). De los ocho géneros foliosos reconocidos, Pyropia es el género con mayor diversidad dentro de la familia, con 74 especies, mientras que Porphyra es el >segundo género con mayor diversidad, con 58 especies aceptadas taxonómicamente (Guiry & Guiry, 2019).
Porphyra y Pyropia han sido registrados en todos los océanos del mundo y han sido incluidos en estudios taxonómicos moleculares y de discriminación de especies de las Bangiales foliosas, tanto a nivel global como regional (ej. Brodie et al., 2008; Mols-Mortensen et al., 2014).
Dichos estudios indican que la simplicidad morfológica que presentan las especies de estos géneros dificulta su identificación.
En Chile, el género Pyropia fue abordado por primera vez con herramientas moleculares por Ramírez et al. (2014), con la descripción de Pyropia orbicularis M. E. Ramírez, L. Contreras Porcia y M.-L. Guillemin, en base al marcador cloroplastidial rbcL (ribulosa 1.5-bifosfato carboxilasa/oxidasa subunidad mayor). Posteriormente, en la costa chilena, Guillemin et al. (2016) estimaron una alta diversidad de especies foliosas de Bangiales en base a rbcL y al marcador mitocondrial COI (citocromo oxidasa I), con la delimitación de ocho especies de Porphyra (Porphyra munfordii, Porphyra sp. CHB, Porphyra sp. CHC, Porphyra sp. CHD, Porphyra sp. CHE, Porphyra sp. CHF, Porphyra sp. FIG y Porphyra sp. FIH) y ocho especies de Pyropia (Pyropia sp. CHG, Pyropia sp. CHH, Pyropia sp. CHI, Pyropia sp. CHJ, Pyropia sp. CHK, Pyropia sp. FIA y Pyropia sp. FID). Ninguna de las especies de Porphyra catalogadas por Ramírez & Santelices (1991) para Chile coincidió con una de las especies determinadas molecularmente, denotando el problema de distinguir las especies en base a morfología y el mal uso de los nombres asignados a especímenes en diferentes partes del mundo (Guillemin et al., 2016). Por el contrario, en el Perú todavía no se ha realizado un estudio profundo de la sistemática molecular de Porphyra y Pyropia. Solo Calderón (2017), al realizar un análisis filogenético de las secuencias rbcL de cinco especímenes foliosos pertenecientes a Bangiaceae, sugirió la presencia de P. munfordii en la costa norte (Salaverry, La Libertad) y dos especies no identificadas de Pyropia, una en la costa norte (Salaverry, La Libertad) y otra en la costa central (Ancón y Callao, Lima). Lamentablemente, estas secuencias no se encuentran depositadas en GenBank.
Las cinco especies de Porphyra registradas para la costa peruana en base a morfología son: P. columbina Montagne, P. umbilicalis (Linnaeus) Kützing, P. pseudolanceolata Krishnamurthy, P. tenera Kjellmae y P. thuretii Dawson (Ramírez & Santelices, 1991). Aún mantienen su estatus genérico a pesar que los análisis filogenéticos realizados con el material tipo (procedente de otras regiones del mundo) indican que estas especies corresponden al género Pyropia, excepto P. umbilicalis que pertenecería al género Porphyra (Sutherland et al., 2011).
Para poder contrastar esta información y poder estimar la verdadera diversidad de estas algas en la costa peruana es necesario realizar análisis moleculares y morfológicos detallados.
El presente trabajo reporta una primera aproximación a la diversidad de Pyropia/Porphyra en el sur del Perú, en el distrito de Marcona, utilizando el marcador mitocondrial COI y el marcador cloroplastidial rbcL. Dentro de estos géneros se comercializan especies para consumo humano directo con el nombre de "cochayuyo", tanto en Marcona como en todo el sur del Perú. Según Acleto (1986), las especies de mayor consumo son las que se conocen hasta ahora como P. columbina y P. pseudolanceolata.
Materiales y métodos
Colecta y preservación de muestras
El muestreo fue realizado durante los años 2016 y 2017, en cuatro localidades del distrito de Marcona (Ica, Perú): Bahía San Nicolás, Playa Siete Huecos, Playa Hermosa y Playa Los Leones (Fig. 1, Tabla 1). El distrito de Marcona presenta 110 km de línea costera, de los cuales 44 están protegidos por ley, correspondiendo la parte norte del distrito a la Reserva Nacional San Fernando y la parte sur, en la Punta San Juan, a la Reserva Nacional Sistema de Islas, Islotes y Puntas Guaneras. La costa de este distrito está formada por grandes extensiones de acantilados y arrecifes que albergan una rica biodiversidad de recursos pesqueros; sus playas ofrecen ensenadas y diversidad de hábitats (IMARPE, 2010)
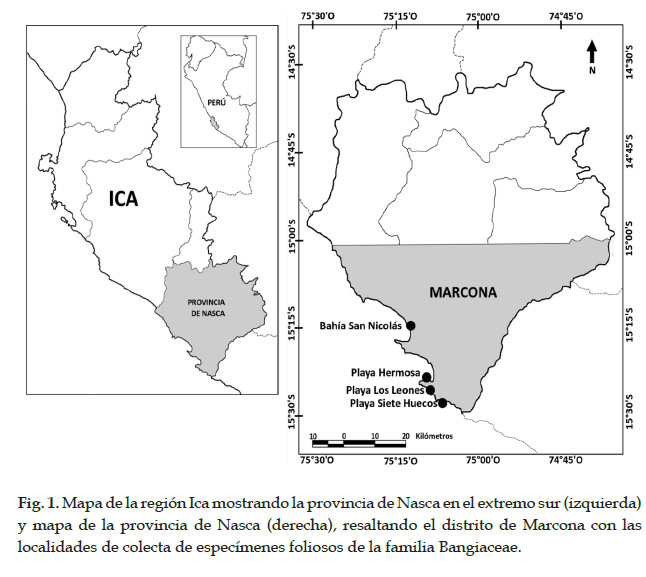
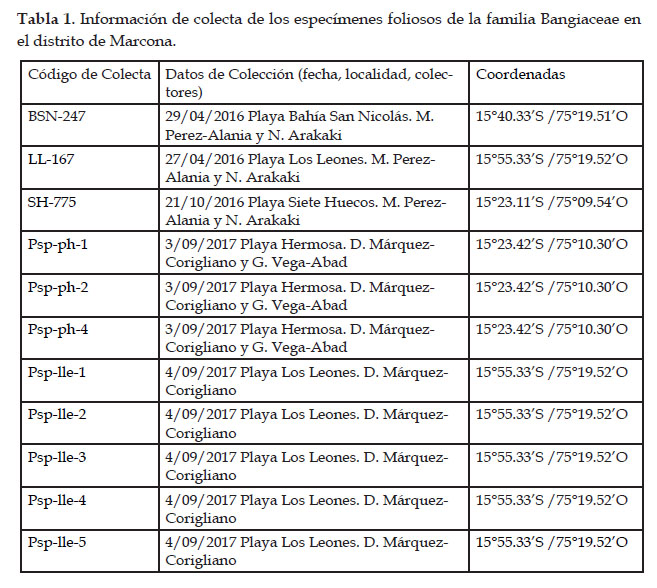
En cada localidad, se colectaron en el intermareal rocoso los especímenes identificados inicialmente como del género Porphyra, en base a su morfología. Se trató de obtener muestras de las distintas morfologías encontradas. Luego de extraer los talos completos desde el disco de fijación, los especímenes colectados fueron trasladados al laboratorio para ser limpiados con agua de mar, separados en base a su morfología y prensados en hojas de herbario. Todos los especímenes seleccionados fueron codificados y fotografiados con una cámara digital o escaneados, antes y después de su herborización. Para el análisis morfológico externo de los especímenes colectados se consideró la forma y las dimensiones del talo, la textura y el color. Una pequeña porción de cada talo fue almacenada en bolsas herméticas con gel de sílica para los análisis moleculares. Los vouchers fueron depositados en la Colección Científica del Instituto del Mar del Perú - IMARPE.
Análisis molecular
El ADN genómico total de las muestras almacenadas fue extraído con el kit GF-1 Nucleic Acid Extraction (Vivantis Technologies Sdn., Selangor, Malasia). Las secuencias parciales para el marcador mitocondrial citocromo c oxidasa subunidad I (COI) fueron obtenidas usando los cebadores GAZF1 y GAZR1, siguiendo el protocolo de amplificación de Saunders (2005). Para el marcador cloroplastidial ribulosa 1.5-bifosfato carboxilasa/oxidasa subunidad mayor (rbcL) se empleó los cebadores F57-R753 y F577-RrbcS, siguiendo los protocolos de Hommersand et al. (1994) y Fredericq & López-Bautista (2003). Todos los productos de PCR fueron purificados y secuenciados por Macrogen Inc. (Seúl, Corea del Sur).
Las secuencias fueron editadas con Codon Code Aligner 2.0 (CodonCode Corporation, MA, Estados Unidos) y alineadas usando el algoritmo ClustalW, implementado en Bioedit 7.0.5 (Hall, 1999). Las secuencias obtenidas fueron analizadas dentro del contexto de especies descritas o identificadas molecularmente como Porphyra y Pyropia y otras Bangiaceae foliosas, utilizando 58 secuencias COI y 82 secuencias rbcL disponibles en las bases de datos de GenBank (Anexo 1). Cada paquete de datos (COI y rbcL) fue evaluado con el programa JModelTest2 (Darriba et al., 2015) para identificar el modelo apropiado de evolución de secuencias en base al Criterio de Información de Akaike (AIC), resultando el modelo GTR+I+G para ambos marcadores. En las reconstrucciones filogenéticas se consideró a Smithora naiadum como grupo externo. Las relaciones filogenéticas fueron inferidas en base al análisis de Máxima Verosimilitud (ML) con el programa RaxML 7.2.6 (Stamatakis, 2014) y 1000 pseudoréplicas bootstraps. El análisis de inferencia bayesiana (IB) fue realizada usando MrBayes 3.2.1 (Huelsenbeck & Ronquist, 2001), con cuatro cadenas de Monte Carlo Markov (MCMC) ejecutadas simultáneamente por cinco millones de generaciones, tomando muestra de un árbol cada 1000 generaciones. El 25% del total de árboles fue descartado y los árboles restantes fueron usados para calcular la topología del árbol consenso y las probabilidades posteriores (PP).
Para aquellos grupos filogenéticos que incluían secuencias de Marcona se estimaron las distancias genéticas, calculando los valores de distancia p (p-distance) intraespecífica e interespecífica usando MEGA v7 (Kumar et al., 2016).
Resultados
Análisis molecular
De los 11 especímenes foliosos de la familia Bangiaceae colectados en Marcona, Perú, se obtuvieron 11 secuencias del marcador plastidial rbcL (875 pares de bases, pb) y ocho secuencias del marcador mitocondrial COI (597 pb). Debido a problemas de amplificación, no se cuenta con secuencia COI para tres especímenes (Tabla 2).
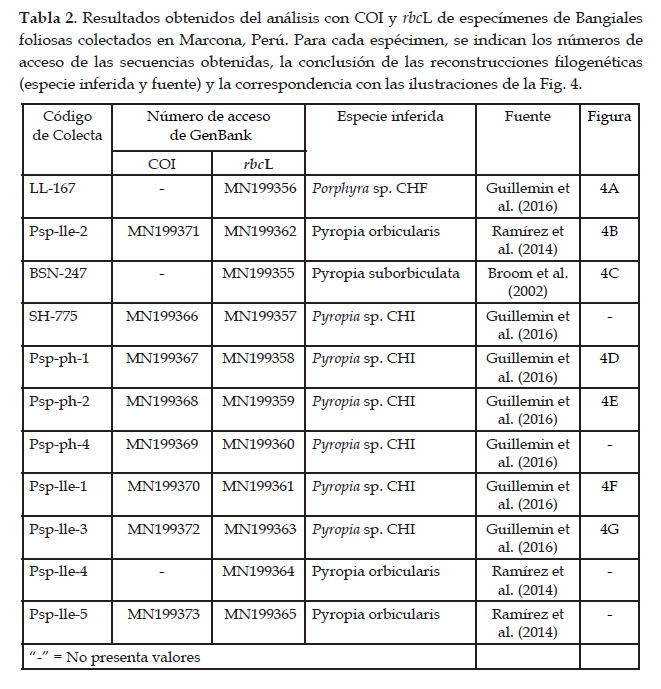
Para ambos marcadores, las topologías de los árboles generados por los análisis IB y ML fueron similares (datos no mostrados). Por otra parte, los valores de soporte de los clados principales fueron comparables entre los análisis, al considerar los soportes superiores a 80 y 70 para IB y ML, respectivamente. En particular, en todos los análisis y marcadores, los clados de Porphyra y de Pyropia estuvieron bien soportados, con valores de soporte superiores a 96 para rbcL y a 83 para COI (Figs 2 y 3). De los especímenes de Marcona, ocho presentaron secuencias COI y rbcL incluidas en el clado de Pyropia. Los especímenes BSN-247 y Psp-Ile-4, aunque secuenciados con rbcL solamente, también pertenecen al clado de Pyropia. Finalmente, un solo espécimen, LL-167, presenta una secuencia rbcL incluida en el clado de Porphyra (no se pudo obtener información para COI).
A nivel específico, se consideraron los clados bien soportados para asignar los especímenes a especies (Tabla 2). La secuencia rbcL del espécimen LL-167 se agrupa con la secuencia de un espécimen de Porphyra sp. CHF procedente de Coquimbo, Chile, con 5 pb de diferencia. La divergencia intraespecífica promedia para Porphyra sp. CHF (secuencias de Perú y Chile) fue de 0.24 (error estándar 0.07).
En el clado Pyropia sp. CHI, se agrupan secuencias de un espécimen de Arica, Chile (frontera con el Perú) y de seis especímenes de Marcona para ambos marcadores (Figs 2 y 3); las localidades comparten haplotipos rbcL pero no COI. La divergencia intraespecífica promedia de Pyropia sp. CHI fue de 0.44 (error estándar 0.12), al considerar las secuencias de Chile y Perú.
Tres especímenes de Marcona presentaron tres secuencias rbcL y dos secuencias COI que se agruparon con la secuencia de Py. orbicularis de Maitencillo, Chile, con valores de soporte mayores a 97 (Figs. 2 y 3); un único haplotipo es compartido entre localidades para rbcL, mientras que se observa una diferencia de 6 pb entre secuencias del Perú y de Chile para COI.
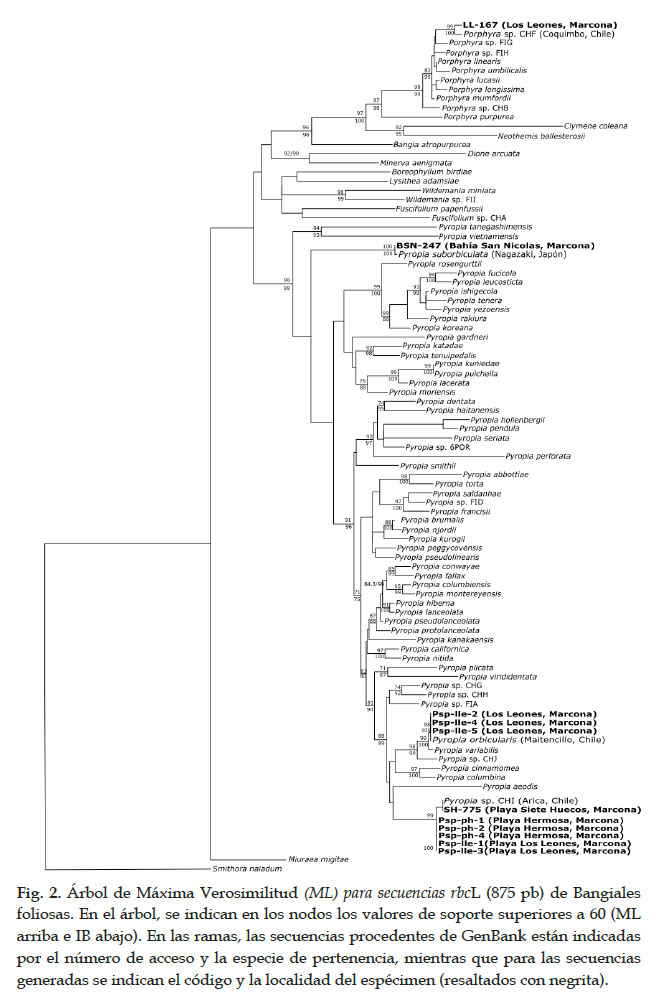
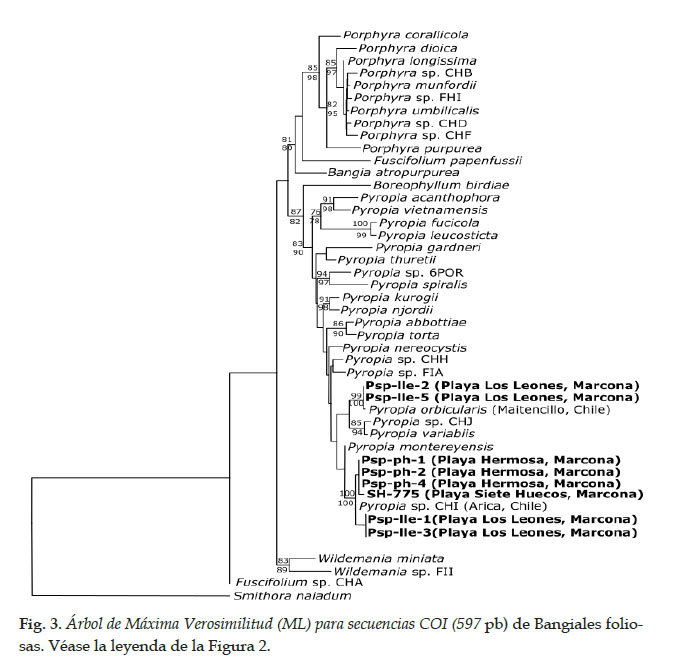
Finalmente, un espécimen con secuencia rbcL se agrupó con la secuencia de Py. suborbiculata de Nagasaki, Japón (Fig. 2), presentando una diferencia de 4 pb.
Análisis morfológico
Los especímenes colectados en Marcona, asociados en base a la filogenia a una especie de Porphyra (Porphyra sp. CHF) y a tres especies de Pyropia (Py. orbicularis, Py. suborbiculata y Pyropia sp. CHI), se presentan en la Fig. 4. El único espécimen de Porphyra sp. CHF presenta una forma rosetada alargada de color verde oscuro con una altura de 3.5 cm y anchura de 5 cm (Fig. 4A), semejante a unas de las formas de Porphyra sp. CHF encontradas en Chile por Guillemin et al. (2016).

Con respecto a las especies de Pyropia, tres especímenes identificados como Py. orbicularis en este estudio presentan una morfología similar entre ellos (Fig. 4B), con una fronda orbicular verde grisácea que varía entre 6 y 11 cm de altura y entre 5 y 9 cm de anchura, semejante al espécimen tipo de esta especie descrito por Ramírez et al. (2014). El espécimen de la especie Py. suborbiculata colectado (Fig. 4C) presenta un talo pequeño (3.5 cm) en forma de corneta de color rosáceo, semejante al espécimen tipo descrito por Kjellman (1896). Finalmente, los seis especímenes de Pyropia sp. CHI colectados en Marcona (playas Hermosa, Los Leones y Siete Huecos) presentan una variedad de morfologías que incluye fronda lanceolada entera (Fig. 4D), fronda laminar entera (Fig. 4E), fronda laminar ondulada con perforaciones (Fig. 4F) y frondas lanceoladas (Fig. 4G); estas morfologías son muy diferentes a las reportadas para especímenes de Chile por Guillemin et al. (2016).
Discusión
Utilizando la evidencia molecular, el presente trabajo reporta la presencia de tres especies de Pyropia y una especie de Porphyra en el extremo sur de la costa central del Perú, sumándose a los esfuerzos actuales en diversas regiones del mundo para caracterizar las Bangiales foliosas. La mayor diversidad específica de Pyropia en Marcona es congruente con lo reportado a nivel mundial (Sutherland et al., 2011).
Para las algas rojas, rbcL es un marcador recomendado para ubicar especies en un contexto filogenético, debido a su evolución más conservada con respecto a otros marcadores (Saunders & Kucera, 2012; Leliaert et al., 2016). Con este marcador se ha reevaluado la identidad de especies de Porphyra y de Pyropia (Broom et al., 2002; Nelson & Broom, 2010) o se han identificado nuevas especies (e.g. Lindstrom et al., 2015; Nelson, 2013). No obstante, a un nivel taxonómico menor, el mayor poder de discriminación del marcador COI permite realizar análisis de delimitación de especies y de filogeografía en algas (e.g. Leliaert et al., 2016; Guillemin et al., 2016; Reddy et al., 2017). Las dificultades de amplificación reportadas aquí para el COI son consistentes con los trabajos que mencionan la dificultad de amplificar COI en algunos grupos de algas rojas, así como el de comparar los especímenes de Porphyra y Pyropia debido a la limitada disponibilidad de secuencias en GenBank (Saunders, 2008; Guillemin et al., 2016; Reddy et al., 2017).
En base a la formación de clados filogenéticos bien soportados, los once especímenes colectados en Marcona, Perú, fueron asignados a especies, todas previamente descritas o reportadas en otras regiones. A continuación se presenta la información de distribución y morfología para las tres especies compartidas entre las costas de Chile y Perú (Py. orbicularis, Pyropia sp. CHI, Porphyra sp. CHF) y para la especie de mayor rango de distribución (Py. suborbiculata).
Pyropia orbicularis
Py. orbicularis fue inicialmente descrita para la costa central de Chile en base a especímenes previamente identificados como "Porphyra columbina" (Ramírez et al., 2014); luego Guillemin et al. (2016) reportaron una distribución geográfica entre 32°S y 53°S en la costa chilena. Con el presente estudio, se extiende el rango de distribución de Py. orbicularis hacia el norte, hasta 15°S, cubriendo así tres provincias biogeográficas. Los especímenes del Perú con sus frondas de forma orbicular corresponden a las características morfológicas indicadas por Ramírez et al. (2014).
Pyropia sp. CHI
Pyropia sp. CHI se encontró en las cuatro localidades consideradas en el presente estudio, siendo la especie de mayor abundancia (6 de los 11 especímenes caracterizados molecularmente). Había sido reportada previamente en la costa norte de Chile, en Arica (18°32’S/70°19’O) y en Coquimbo (30°01’S/71°23’O; Guillemin et al., 2016), por lo que su rango geográfico se extiende al menos entre 15°S y 30°S. La divergencia intraespecífica promedia de Pyropia sp. CHI fue inferior a los valores presentados por Reddy et al. (2018), confirmando una sola entidad. No obstante, la gran diversidad de morfologías detectada en Marcona contrasta con lo reportado en Chile, aspecto que debería ser profundizado en futuras investigaciones.
Porphyra sp. CHF
Porphyra sp. CHF ha sido reportada en Chile, desde la Región de Antofagasta (23°46’S/70°28’O) hasta la Región del Biobío (36°31’°S/72°57’O, Guillemin et al., 2016) y el presente trabajo extiende su distribución hasta 15°S, al reportar la presencia de un espécimen en Marcona. La divergencia intraespecífica obtenida para esta especie fue inferior al valor de distancia p máxima de las especies de Porphyra para rbcL presentada por Reddy et al. (2018), corroborando una sola entidad. La plasticidad morfológica en Porphyra sp. CHF de Chile es bastante notoria, con una diversidad de formas (Guillemin et al., 2016), lo que no ha podido ser corroborado en Marcona, al contar con solo un espécimen secuenciado.
Pyropia suborbiculata
Pyropia suborbiculata es la única especie de Bangiales foliosas conocida hasta la fecha que se encuentra distribuida en ambos hemisferios (Nelson et al., 2005). Inicialmente fue descrita en Nagasaki, Japón por Kjellman (1897) y luego reportada en varias costas de los países asiáticos (Corea, China y Vietnam) (Aguilar-Rosas & Aguilar-Rosas, 2003). Posteriormente, bajo la evidencia molecular se confirmó la presencia de Py. suborbiculata en Nueva Zelanda, Australia, Brasil y el Pacífico mexicano (Milstein et al., 2012), en el Atlántico norteamericano (Broom et al., 2002, Aguilar-Rosas & Aguilar-Rosas, 2003) y en la Península Ibérica (Vergés et al., 2013). Probablemente, esta amplia distribución sea el resultado de múltiples introducciones y colonizaciones exitosas (Broom et al., 2002; Verges et al., 2013). El espécimen de Marcona identificado como Py. suborbiculata presenta una fronda entera pequeña (4 cm) de forma cónica, lo que corresponde a las características mencionadas por Broom et al. (2002) para el espécimen tipo de Nagasaki, Japón. El reporte de esta especie en el Perú representa el primer registro para la costa del Pacífico sudamericano.
Diversidad de especies de Bangiales foliosas en el Perú
Si bien el muestreo no pretende ser representativo de la diversidad específica local de los géneros Porphyra y Pyropia, cabe destacar que Marcona es una región de colecta importante de "cochayuyo". Aun cuando Acleto (1986) indica que bajo este nombre las especies de mayor consumo humano en el Perú son P. columbina y P. pseudolanceolata (ahora transferidas al género Pyropia), ninguna ha sido encontrada en el presente estudio. Tampoco se encontró evidencia de la presencia de P. tenera y P. thuretti (ahora en el género Pyropia) ni de P. umbilicalis, especies reportadas para Perú sobre la base de caracteres morfológicos. Finalmente, P. monfordii, reportada por Calderón (2017) en base a datos moleculares en el norte del Perú, no fue encontrada.
Un estudio más exhaustivo, cubriendo mejor la costa peruana y con un mayor número de especímenes, es necesario para corroborar qué especies de los géneros Porphyra y Pyropia se encuentran distribuidas en Perú y en particular cuáles son las más frecuentemente consumidas. Esta información taxonómica y de delimitación de especies es crucial al momento de establecer planes de manejo y conservación de los recursos algales (e.g. por revisión Tellier et al., 2011).
Conclusiones
Se muestra la presencia en la costa peruana de tres especies de Pyropia y una especie de Porphyra, que no corresponden a las especies descritas para el país en base a caracteres morfológicos. Tres de las especies encontradas, Porphyra sp. CHF, Py. orbicularis y Pyropia sp. CHI, tienen un rango de distribución por ahora limitado a Chile y Perú. Py. suborbiculata, si bien es de amplia distribución mundial, no había sido reportada previamente en la costa del Pacífico sudamericano.
Agradecimientos
Agradecemos el apoyo de Melissa Perez Alania y Gustavo Vega Abad en la colecta de muestras y a Karla Pérez-Araneda en la secuenciación de algunas muestras.
Este trabajo fue financiado por el Convenio de Cofinanciamiento 017-2016-FONDECYT - Perú. Las salidas de campo en el año 2016 fueron financiadas por el Convenio de Subvención 129-2015-FONDECYT Perú en Alianza UNALM - IMARPE. Además, este trabajo recibió aporte adicional de la Universidad Católica de la Santísima Concepción (Fondo FAA de la DIN).
Contribución de los autores
La contribución de los autores fue la siguiente: D.M.C., N.A. y P.G.K. participaron en la concepción y diseño del trabajo, D.M.C. y N.A. realizaron el muestreo. D.M.C. lideró la obtención y el análisis de datos genéticos y morfológicos y elaboró el manuscrito. F.T., N.A. y P.G.K. contribuyeron a la interpretación de los datos y con revisiones críticas del manuscrito. Todos los autores han leído el manuscrito final y aprobado la revisión.
Conflicto de intereses
Todos los autores aprueban la versión enviada de este manuscrito y declaran no tener conflictos de intereses.
Literatura citada
Acleto, C. 1986. Algas marinas del Perú de importancia económica. Museo de Historia Natural "Javier Prado" UNMSM. 107 p (Serie de divulgación N° 5). [ Links ]
Aguilar-Rosas, R. & L. E. Aguilar-Rosas. 2003. El género Porphyra (Bangiaceae, Rhodophyta) en la costa Pacífico de México. I. Porphyra suborbiculata Kjellman. Hidrobiológica 13(1): 51-56. [ Links ]
Brodie, J.; A. Mols-Mortensen; M. E. Ramirez; S. Russell & B. Rinkel. 2008. Making the links: Towards a global taxonomy for the red algal genus Porphyra (Bangiales, Rhodophyta). Journal of Applied Phycology 20(5): 939-949. DOI 10.1007/s10811-008-9315-7 [ Links ]
Broom, J. E.; W. A. Nelson; C. Yarish; W. A. Jones; R. Aguilar-Rosas & L. Aguilar-Rosas. 2002. A reassessment of the taxonomic status of Porphyra suborbiculata, Porphyra carolinensis and Porphyra lilliputiana (Bangiales, Rhodophyta) based on molecular and morphological data. European Journal of Phycology 37: 227-235. DOI: 10.1017/S0967026202003566 [ Links ]
Calderón, M. 2017. Filogenia molecular de algunas algas marinas rojas del Perú basada en análisis de ADN plastidial. Facultad de Ciencias Biológicas. Universidad Nacional Mayor de San Marcos. 275 p. [ Links ]
Fredericq, S. & J. López-Bautista. 2002. Characterization and phylogenetic position of the red alga Besa papillaeformis Setchell: an example of progenetic heterochrony?. Constancea 83: 1-12. [ Links ]
Guillemin, M. L.; L. Contreras-Porcia; M. E. Ramírez; E. C. Macaya; C. B. Contador; H. Woods; C. Wyatt & J. Brodie. 2016. The bladed Bangiales (Rhodophyta) of the South Eastern Pacific: Molecular species delimitation reveals extensive diversity. Molecular Phylogenetics and Evolution 94: 814-826. DOI: 10.1016/j.ympev.2015.09.027. [ Links ]
Guiry, M. D. & G. M. Guiry. 2019. AlgaeBase. World- wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; consultado el 18 marzo de 2019. [ Links ]
Hall, T. 1999. Bioedit: A user-friendly biological sequence alignment editor and analysis program for Windows 95. Nucleic Acids Symposium Series 41: 95-98. [ Links ]
Hommersand, M. H.; S. Fredericq & D. W. Freshwater. 1994. Phylogenetic systematics and biogeography of the Gigartinaceae (Gigartianles, Rhodophyta) based on sequence analysis of rbcL. Botanica Marina 37: 193-203. [ Links ]
IMARPE. 2010. Caracterización bio-oceanográfica del área marino costera entre Punta San Juan y Playa Yanyarina. Informe del Instituto del Mar del Perú 42(3): 282-294. [ Links ]
Kjellman, F. R. 1897. Japanska arter af slägtet Porphyra. Bihang Till K. Svenska Vet-Akad. Handlingar 23: 1±34. [ Links ]
Kucera, H. & G. W. Saunders. 2012. A survey of Bangiales (Rhodophyta) based on multiple molecular markers reveals cryptic diversity. Journal of Phycology 48(4): 869-882. DOI: 10.1111/j.1529-8817.2012.01193.x. [ Links ]
Kumar, S.; G. Stecher & K. Tamura. 2016. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Molecular Biology and Evolution 33(7): 1870-1874. DOI: 10.1093/molbev/msw054. [ Links ]
Leliaert, F.; H. Verbruggen; P. Vanormelingen; F. Steen; J. M. López-Bautista; G. C. Zuccarello & O. De Clerck. 2014. DNA-based species delimitation in algae. European Journal of Phycology 49(2): 179-196. DOI: 10.1080/09670262.2014.904524. [ Links ]
Lindstrom, S. C.; M. R. Lindeberg & D. A. Guthrie. 2015. Marine macroalgae of the Aleutian Islands: I. Bangiales. Algae 30(4): 247-263. DOI: 10.4490/algae.2015.30.4.247 [ Links ]
Milstein, D.; A. S. Medeiros; E. U. Oliveira & M. C. Oliveira. 2015. Native or Introduced? A re-evaluation of Pyropia species (Bangiales, Rhodophyta) from Brazil based on molecular analyses. European Journal of Phycology 50: 37- 35. DOI: 10.1080/09670262.2014.982202. [ Links ]
Mols-Mortensen, A.; C. D. Neefus; P. M. Pedersen & J. Brodie. 2014. Diversity and distribution of foliose Bangiales (Rhodophyta) in West Greenland: A link between the North Atlantic and North Pacific. European Journal of Phycology 49(1): 1-10. DOI: 10.1080/09670262.2013.871062 [ Links ]
Nelson, W. A. & J. E. Broom. 2010. The identity of Porphyra columbina (Bangiales, Rhodophyta) originally described from the New Zealand subantarctic islands. Australian Systematic Botany 23(1): 16-26. DOI: 10.1071/SB09032. [ Links ]
Nelson, W. A.; T. J. Farr & J. E. Broom. 2005. Dione and Minerva, two new genera from New Zealand circumscribed for basal taxa in the Bangiales. Phycologia 44(2): 139-145. DOI: 10.2216/0031-8884(2005)44[139:DAMTNG]2.0.CO;2. [ Links ]
Ramírez, M. E. & B. Santelices. 1991. Catálogo de las algas marinas bentónicas de la costa temperada del Pacífico de Sudamérica. Monografías Biológicas 5: 1-437. [ Links ]
Ramírez, M. E.; L. Contreras-Porcia; M. L. Guillemin; J. Brodie; C. Valdivia; M. R. Flores-Molina; A. Núñez; C. B. Contador & C. Lovazzano. 2014. Pyropia orbicularis sp. nov. (Rhodophyta, Bangiaceae) based on a population previously known as Porphyra columbina from the central coast of Chile. Phytotaxa 158(2): 133-153. DOI: 10.11646/phytotaxa.158.2.2. [ Links ]
Reddy, M; O. De Clerck; F. Leliaert; R. A. Anderson & J. J. Bolton. 2017. A rosette by any other name: Species diversity in the Bangiales (Rhodophyta) along the South African coast. European Journal of Phycology 53(1): 67-82. DOI: 10.1080/09670262.2017.1376256. [ Links ]
Sánchez, N.; A. Vergés; C. Peteiro; J. E. Sutherland & J. Brodie. 2014. Diversity of bladed Bangiales (Rhodophyta) in Western Mediterranean: Recognition of the genus Themis and descriptions of T. ballesterosii sp. nov., T. iberica sp. nov., and Pyropia parva sp. nov. Journal of Phycology 50(5): 908-929. DOI: 10.1111/jpy.12223. [ Links ]
Stamatakis, A. 2014. RAxML v8: A tool for phylogenetics analysis and post-analysis of large phylogenies. Bioinformatics 30(9): 1312-1313. DOI: 10.1093/ bioinformatics/btu033. [ Links ]
Sutherland, J. E.; S. C. Lindstrom; W. A. Nelson; J. Brodie; M. D. Lynch; M. S. Hwang; H. G. Choi; M. Miyata; N. Kikuchi; M. C. Oliveira; T. Farr; C. Neefus; A. Mols-Mortensen; D. Milstein & K. M. Müller. 2011. A new look at an ancient order: Generic revision of the Bangiales (Rhodophyta). Journal of Phycology 47(5): 1131-1151. DOI: 10.1111/j.1529-8817.2011.01052.x. [ Links ]
Tellier, F., J. M. A. Vega, B. R. Broitman, J. A. Vásquez, M. Valero, & S. Faugeron. 2011. The importance of having two species instead of one in kelp management: the Lessonia nigrescens species complex. Cahiers de Biologie Marine 52: 455-465. [ Links ]
Autor para correspondencia:
narakaki@imarpe.gob.pe
Recibido: 11-IV-2019
Aceptado: 17-V-2019
Publicado online: 15-VIII-2019
Publicado impreso: 31-VIII-2019

