











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Los bosques húmedos montanos y premontanos de los Andes Tropicales son unos de los ecosistemas más diversos del mundo en lo que se refiere al número de especies (Krömer, Gradstein & Aceby 2007), luego de las selvas Amazónicas. En este contexto, las especies de flora epífita vascular de los taxa botánicos Orchidaceae, Araceae, Bromeliaceae y Pteridophytas son un componente crucial para la conservación de otras formas de vida de estos bosques, incluyendo el bosque mismo, por brindar el servicio ecosistémico de captar el recurso hídrico de la humedad ambiental (Calaway H Dodson, 2003; Krömer et al., 2007). Estas plantas crean un microclima y microhábitat idóneo para especies de invertebrados que sirven de alimento a redes tróficas de vertebrados. Actualmente, los bosques húmedos están desapareciendo rápidamente como resultado de la creciente presión antrópica que soportan. Aunque que son reconocidos por su diversidad y endemicidad, la cantidad de información sobre las orquídeas de estos bosques es limitada, así como su ecología, diversidad alfa y beta altitudinal en gradientes.
Las orquídeas constituyen una numerosa familia de plantas con flores, las Angiospermas. Se estima que la familia Orchidaceae existe desde hace 120 millones de años, siendo una de las familias más evolucionadas, con un alto grado de diversidad (Calaway H Dodson, 2003; Freuler, 2007). Cuenta con alrededor de 25,000 especies descritas, repartidas en 750 géneros (MINAM, 2015b). En los Andes Tropicales, la mayor diversidad observada de orquídeas se da sobre los 2000 m.s.n.m. (Calaway H Dodson, 2003), aunque esto varía entre los géneros de orquídeas. Según los estudios de Bennett & Christenson (1993, 1995) el Perú alberga alrededor de 3,000 especies de Orchidaceae, ocupando toda su extensión territorial continental.
Asimismo, la familia Orchidaceae constituye uno de los grupos de plantas más vulnerables a los cambios ambientales, debido a su alto grado de asociación a microambientes específicos, además de ser indicadoras de calidad de hábitat, presencia o abundancia de otras especies, entre otros (Krömer et al., 2007). Una amenaza adicional es la explotación de especies para su comercialización. Las orquídeas son plantas muy vistosas y apreciadas por su valor ornamental, lo que ha ocasionado que exista una fuerte presión de extracción, debido a la demanda del comercio nacional e internacional, poniendo en peligro a muchas especies. Esta situación ha ocasionado que todas las orquídeas sean incluidas en la lista CITES, que es la Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestre, donde se registra a las especies cuyo comercio debe ser regulado para prevenir su extinción, en otras palabras, exterminación (MINAM, 2015a).
Dentro de los estudios publicados sobre listas de especies de orquídeas andinas, la mayoría amplía la distribución de taxones conocidos, incluyendo nuevos reportes para Perú. No obstante, estos estudios han sido realizados principalmente en regiones más sureñas, como La Libertad (Rodríguez, Alvítez, Pollack, et al., 2014), Cusco (Moscoso, Salinas, & Nauray 2003), o Tingo María (Ocupa, Díaz, Yupanqui, et al., 2017) y escasos en el noroeste de Perú. Esto ha generado un vacío de información para las especies de bosques nublados en la zona biogeográfica Depresión de Huancabamba, en la frontera Perú Ecuador. Esta investigación tuvo como objetivo determinar la diversidad, distribución altitudinal, distribución vertical y estado fenológico de las especies de Orchidaceae presentes en el lado occidental de la quebrada Colorada, distrito de El Carmen de la Frontera, provincia de Huancabamba, departamento de Piura, norte de Perú.
Materiales y métodos
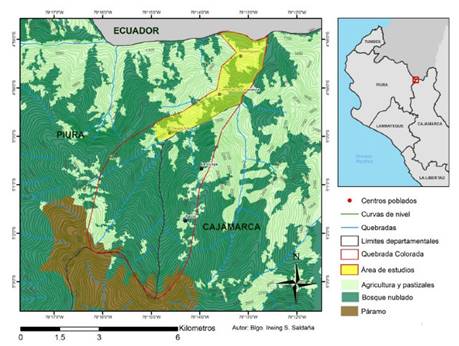
Área de estudios
El área de estudios se localiza en el ámbito de la quebrada Colorada, distrito El Carmen de la Frontera, provincia de Huancabamba, departamento de Piura. Esta quebrada divide los departamentos de Piura y Cajamarca, y a su vez es afluente del río Blanco, límite de ese sector de la frontera Perú-Ecuador. La quebrada Colorada recorre el rango altitudinal desde los 950 m.s.n.m. en la zona baja del pueblo El Hormiguero, límite geopolítico con Ecuador, hasta los 3,600 m.s.n.m. en el límite con el Santuario nacional TabaconasNamballe. Dentro de este contexto, se delimitó el área de estudios entre los 1,100 y 2,100 m.s.n.m.
De las 2,702 has la microcuenca quebrada Colorada, la cobertura del suelo (Fig. 1) es del 71,65% de bosques (1936 has), 24,46% de áreas de agricultura/pastizales (661 has) y 3,89% de páramos (105 has) (MINAM, 2015c).
Diseño de estudio
Se realizó una salida preliminar al área de estudio para identificar los rangos máximos y mínimos que podían ser evaluados dependiendo del acceso y rutas disponibles. Posteriormente, se estratificó el área de estudios respetando los límites altitudinales de las zonas de vida presentes, de modo que se generaron tres estratos altitudinales. E1: de 1,100 a 1,300 m.s.n.m., con bosque húmedo premontano bajo Tropical, E2: de 1,300 a 1,500 m.s.n.m., con bosque húmedo premontano Tropical, y E3: de 1,500 a 2,200 m.s.n.m., con bosque húmedo montano bajo Tropical (MINAM, 2015c). Se realizaron tres salidas de estudio, evaluación I: julio 2016, evaluación II: diciembre 2016, evaluación III: agosto 2017, visitando todos los estratos en cada visita. En estrato, se ejecutaron tres parcelas de 50 x 50 m dentro de cada estrato. En cada parcela evaluada, se realizó búsqueda intensiva de ejemplares en sustrato rocoso, terrestre y epífito. De este modo, el esfuerzo de muestro fue de 27 parcelas, con 6,75 hectáreas de terreno cubierto (0,24% de la quebrada Colorada). Se colectaron especies litófitas, terrestres y epífitas; estas últimas se buscaron hasta una altura aproximada de 2m, abarcando la primera y segunda zona de clasificación vertical de un árbol o fitóforo (Johansson, 1974). Esta metodología permite colectar el 50% del total de la diversidad para un forofito (árbol hospedero), lo cual permite obtener una muestra regularmente representativa del dosel.
Para la identificación correcta de las especies se emplearon las colectas de campo y fotografías de las plantas y floraciones in situ. Estas fueron contrastadas con la bibliografía disponible y mediante consulta a expertos, como la autoridad mundial en el género Epidendrum Eric Hágsater del Herbario AMO Asociación Mexicana de Orquideología, Instituto Chinoin - México, el especialista en la subfamilia Orchidoideae Gerardo A. Salazar, Universidad Autónoma de México, Günther Gerlach del Jardín Botánico Nymphenburg de Múnich Alemania para el grupo Stanhopeinae y Mark Wilson del Departamento de Biología y Ecología de Organismos, Colorado College, Colorado Springs - EE.UU para el grupo Pleurothallidinae; quienes fueron contactados y se les envió la evidencia fotográfica de los ejemplares de taxonomía compleja para solicitar su identificación. En gabinete, se calculó la densidad poblacional y la frecuencia de cada una de las especies registradas durante las tres salidas de estudio.
Aspectos éticos
No se colectaron especímenes de todas las especies. Las colectas botánicas se realizaron bajo el permiso de colecta como resolución de dirección general N° 247-2016-SERFOR/DGGSPFFS emitida
el 19 de julio de 2016, con ampliación para la zona de estudio mencionada como Quebrada Colorada (694152.99 9447200.99 UTM) mediante resolución de dirección general N° 430-2017-SERFOR/ DGGSPFFS, autorizado por el ente estatal SERFOR "Servicio Nacional Forestal y de Fauna Silvestre", encargado de brindar los permisos de investigación y colecta en áreas nacionales no protegidas. El permiso fue gestionado y concedido al equipo de la Universidad Nacional Pedro Ruiz Gallo, dentro del proyecto de "Estudios Taxonómicos selectos de la flora del norte del Perú", al cual pertenece el investigador LOH.
Resultados
Se registraron 30 especies y 18 taxa identificados a nivel de género (Tabla 1), distribuidos en 26 géneros, de los cuales Epidendrum (con 8 taxa) y Pleurothallis (con 5 taxa) fueron los más diversos. Los demás géneros estuvieron representados por 1 (15 géneros) o 2 taxa (8 géneros). Adicionalmente, se realizaron dos nuevos reportes de Orchidaceae para Perú: Maxillaria pergracilis (Figura 5D) y Sarcoglottis grandiflora (Figura 3E).
Tabla 1 Riqueza de especies de Orchidaceae por estrato altitudinal en el área de estudios. E1: 1100-1300 m.s.n.m., E2: 13001500 m.s.n.m., E3: 1500-2200 m.s.n.m. *: nuevo registro para Perú, **: especie endémica de Perú.
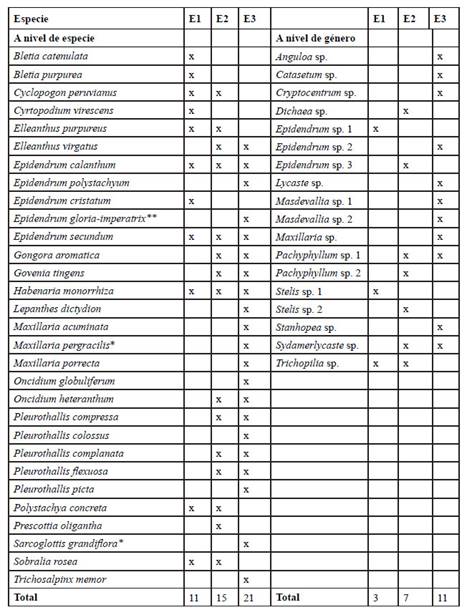

Fig. 3 A. Epidendrum calanthum, B. Epidendrum polystachyum, C. Epidendrum cristatum, D. Epidendrum gloria-imperatrix, E. Epidendrum secundum, F. Gongora aromatica.

Fig. 5 A. Oncidium heteranthum, B. Pleurothallis compressa, C. Pleurothallis complanata, D. Pleurothallis flexuosa, E. Pleurothallis picta, F. Polystachya concreta.
Se reportaron cuatro especies exclusivas del estrato E1 (1100-1300 m.s.n.m.): Bletia catenulata, B. purpurea, Cyrtopodium virescens y Epidendrum cristatum; ninguna exclusiva del estrato E2 (1300-1500 m.s.n.m.); y 11 exclusivas del estrato E3 (1500-2200 m.s.n.m.): E. polystachyum, E. gloriaimperatrix, Lepanthes dictydion, Maxillaria acuminata, Maxillaria pergracilis, Maxillaria porrecta, Oncidium globuliferum, Pleurothallis colossus, P. picta, Sarcoglottis grandiflora y Trichosalpinx memor. Además, cinco especies se distribuyeron a lo largo de todo el rango altitudinal del área de estudios: Elleanthus virgatus, Epidendrum calanthum, E. secundum, Habenaria monorrhiza y Polystachya concreta (Tabla 1).
El periodo de mayor floración reportado se dio en el mes de agosto, con 16 especies/taxa en floración, versus 10 en julio y 12 en diciembre. Dos especies fueron reconocida a nivel de especie empleando vestigios florales, además de características taxonómicas únicas de su parte vegetativa para el reconocimiento, Pleurothallis colossus y Elleanthus virgatus. Solo una especie floreció durante todo el periodo evaluado, Epidendrum secundum (Tabla 2).
Tabla 2 Estados fenológicos y hábito de las especies de Orchidaceae reportadas en el estudio. Estados fenológicos: gris oscuro: floración, gris claro: vegetativo, X: vestigios de floración que aportaron suficiente evidencia para determinar la especie, blanco: no reportada durante ese periodo. Hábito: T1: terrestre umbrófila, T2: terrestre en hábitats abiertos, L: litófita, E: epífita.
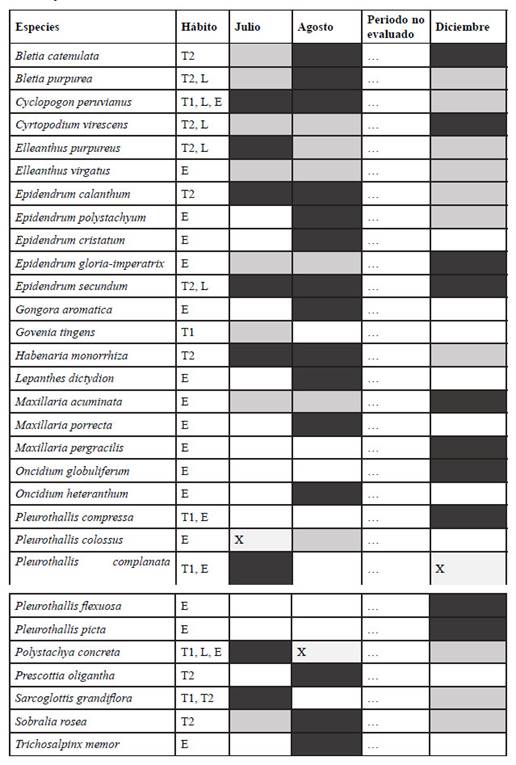
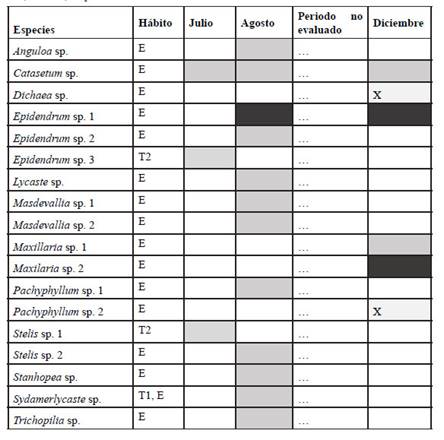
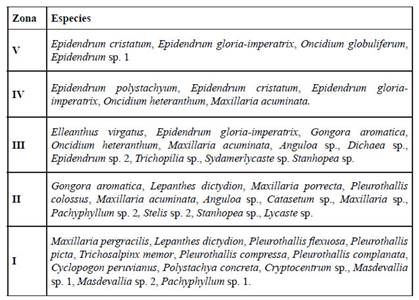
Se reportaron 29 especies exclusivamente epífitas, una especie exclusivamente terrestre umbrófila, siete especies exclusivamente terrestres de hábitats abiertos y ninguna exclusivamente litófila. Dos especies, Cyclopogon peruvianus y Polystachya concreta fueron encontradas en todos los hábitos, considerando solo un tipo de hábito terreste (Tabla 1, 3). Las especies de hábito epífito se distribuyeron entre todas las zonas de Johansson (1974), siendo la zona más diversa la I, con 13 especies/ taxa, seguida de II con 12 especies/taxa, III, con 11 especies/taxa, IV, con 5 especies/ taxa y V, con 4 especies/taxa (Tabla 4). Todas las zonas compartieron especies entre sí, encontrando 12 especies exclusivas de la zona I, 7 de la zona II, 4 de la zona III, 1 de la zona IV y 2 de la zona V.
Tabla 3 Estados fenológicos y hábito de las taxa de Orchidaceae reconocidas a nivel de género, reportadas en el estudio. Estados fenológicos: gris oscuro: floración, gris claro: vegetativo, X: vestigios de floración que aportaron suficiente evidencia para determinar la especie, blanco: no reportada durante ese periodo. Hábito: T1: terrestre umbrófila, T2: terrestre en hábitats abiertos, L: litófita, E: epífita.
Discusión
La distribución altitudinal de las especies de orquídeas indicaría que, a mayor altitud, mayor diversidad de orquídeas (Tabla 1). Sin embargo, existe otro factor juega un rol importante y es necesario mencionarlo para poder interpretar adecuadamente esta premisa, la deforestación de los bosques nublados para apertura de pastos y áreas de cultivo. Los impactos antrópicos en la microcuenca tienden a ocupar las altitudes más bajas por la preferencia de cultivos termófilos (Figura 1). En el área de estudios, principalmente se siembra café, siendo este sembrado de manera "bajo sol directo", por lo que los pobladores deforestan por completo sus parcelas antes de sembrarlas. Se reportan también cultivos de café hasta los 2100 m.s.n.m., pero la mayor cantidad de parcelas se ubican bajo los 1500 m.s.n.m. Esto propicia la pérdida de diversidad de especies orquídeas epífitas, principalmente umbrófilas, en el rango de 1100 a 1500 m.s.n.m, algo bastante común en los Andes Tropicales.
Las diferencias entre la diversidad de epífitas encontradas entre los bosques montanos del norte de Perú, pueden explicarse debido a las características particulares del sitio. La historia de uso, el tipo de intensidad de las perturbaciones pasadas, la topografía del lugar y la fauna existente son factores claves en el desarrollo de la dinámica forestal, como la formación de claros, regeneración natural, dispersión, entre otros, y verse reflejados en características del bosque como la riqueza y la composición florística (RasalSánchez, 2012). Más aún, la reducción puntual de la diversidad de epífitas está íntimamente relacionada con estructura de los fitóforos, los árboles que sirven de sustrato a las orquídeas. Bosques con árboles jóvenes presentan menor diversidad de orquídeas debido a una falta de capa de musgo establecida previamente antes del intento de colonización, por lo que este microclima resulta más seco de lo que las orquídeas necesitan (Krömer et al., 2007). Lo anteriormente mencionado y la disponibilidad de luz adecuada son los principales factores microclimáticos para el establecimiento de las orquídeas en los forofitos (Riofrío, Naranjo, Iriondo, et al., 2007).
En este sentido, las 29 especies de orquídeas que son exclusivamente epífitas en el área de estudio se reparten entre las zonas verticales de los fitóforos en relación con la necesidad de luz y humedad idónea para ellas. La mayor cantidad de especies epífitas fueron umbrófilas (n=13), por la preferencia de la zona más baja (zona I, Tabla 4), que recibe menor incidencia de luz y viento. Además, la mayoría de estas trece especies epífitas umbrófilas son miniatura. Esto explica que fisiológicamente no soportan vientos, temperatura y radiación mayor de la que el sotobosque recibe, las zonas de Johansson (1974) más altas resultan letales para ellas. De igual modo, la tala selectiva y apertura de claros en los bosques tienen el mismo efecto sobre ellas. Adicionalmente, es en este estrato (I) en el que se registró dos de los tres nuevos reportes de especies para el Perú, Cryptocentrum pergracile y Pleurothallis complanata.
Los nuevos reportes para Perú, Pleurothallis complanata, Sarcoglottis grandiflora y Maxillaria pergracilis, pueden ser considerados como nuevo reporte o primer reporte documentado de las especies, debido a que algunos autores mencionan su presencia sin ningún dato adicional sobre el registro. S. grandiflora se encuentra desde Entre Ecuador y Argentina, no obstante, ninguna fuente bibliográfica de las orquídeas peruanas lo menciona (Becerra 2007; Bennett & Christenson 1993, 1995; Dodson & Bennett 1989; MINAM 2012; Ocupa et al. 2017; Pupulin 2012; Roque & León 2013; Salinas & Nauray 2016; Schweinfurth 1960, 1961, 1970, 1959), a excepción del libro Orchids Species of Peru (Zelenko & Bermudez 2009) quien solo coloca una mención ligera de la especie, sin detalle. El registro de esta especie era esperado dado su amplio rango de distribución. Pleurothallis complanata está documentada solamente para el área de Morona-Santiago en el sureste de Ecuador (Luer, 2005). Esta área se encuentra en el límite con el departamento de Loreto, justo al oeste de la Cordillera del Cóndor. Probablemente el rango de la especie se encuentre por el sur hasta el río Chinchipe, al cual es afluente la quebrada Colorada donde se realizó el presente estudio. Cryptocentrum pergracile se encuentra entre Colombia y Ecuador en pie de monte entre los 1000 y 1500 m.s.n.m, en el área de estudios se le reportó entre los 1500 y 2200 m.s.n.m (Dodson & Bennett 1989). Las características morfológicas coindicen con la especie, pero la preferencia de hábitat puede deberse a que este sería su extremo sur de distribución, o a que sea una nueva especie afín a M. pergracilis. Nuevas investigaciones y colecta de especímenes son requeridas para confirmar el estatus.
La posible nueva especie afin a Pleurothallis compressa, de la sección Macrophyllae-Fasciculatae, fue reportada con una población regularmente abundante sobre los 2100 m.s.n.m., en bosques pristinos. P. compressa es endémica de Costa Rica, pero no sería el primer caso de especies cuya presencia hace saltos latitudinales entre Centro América y algún país Sudamericano, algunas especies de Epidendrum y Lepanthes tienen este patrón (Dodson & Bennett 1989). Adicionalmente, existen especies Centroamericanas que tienen contrapartes afines filogenética y morfológicamente en los Andes Tropicales, como lo menciona Gentry (1982), por lo que es probable que se trate de una nueva especie no descrita. P. compressa se diferencia de afines dentro de la sacción Macrophyllae-Fasciculatae por presentar ramicaules lateralmente compresos, los costados del sépalo dorsal, sinsépalo, así como los pétalos, reflexos. Los especímenes fotografiados presentaron estas características excepto por el labelo, el cual es el doble de largo (7 mm) que en P. compressa. No se realizaron colectas de especímenes debido a que no se obtuvo permiso de colecta de parte del SERFOR para la fecha del muestreo, y cuando se buscaron poblaciones para la siguiente evaluación, ya no se encontraron en floración. Mayor investigación es requerida para confirmar su estatus y/o describirla.
La única especie endémica de Perú registrada, Epidendrum gloria-imperatrix, reportada en Catamarca, provincia de San Ignacio, Cajamarca (Hágsater, Ayala, Saldaña, et al., 2007), representa un nuevo reporte para Piura (MINAM, 2012). Esta especie es probablemente endémica del valle del río Chinchipe y afluentes, por lo que es esperable que próximamente sea reportada cruzando el límite internacional en Ecuador.
Una evaluación más minuciosa puede aumentar considerablemente y enriquecer el listado de especies reportadas durante el presente estudio, incluyendo así quizás especies nuevas para la ciencia o reportes nuevos para el Perú.




