











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
El cambio climático es una de las más severas preocupaciones para las políticas de los gobiernos y agricultores en las últimas 2 décadas. El cambio en las condiciones climáticas está directamente relacionado con el aumento poblacional, lo cual a su vez crea la necesidad de mantener una alta productividad en los campos de cultivo y la seguridad alimentaria para estas poblaciones (Singh et al., 2009). Sin embargo, esto lleva a los agricultores a utilizar grandes cantidades de fertilizantes químicos y pesticidas para aumentar los rendimientos de los cultivos.
Aquellos fertilizantes son sintetizados químicamente a partir de nitrógeno, fósforo y potasio principalmente los cuales al ser usados indiscriminadamente provocan contaminación del suelo, aire y agua ya sea directa o indirectamente (Galloway et al., 2019) (Youssef & Eissa, 2014). El continuo uso de fertilizantes, biocidas y pesticidas afectan negativamente la microflora natural la cual incluye bacterias, hongos, micorrizas, cianobacterias y protozoarios presentes en la rizósfera causando un desbalance en el ecosistema natural, afectando la promoción del crecimiento, control de enfermedades, tolerancia tanto a sales como al estrés por metales pesados (Dash et al., 2017) (Dash et al., 2018) (Wani et al., 2016). La rizósfera de las plantas es una fina capa de suelo adherida a la superficie de las raíces, allí es donde se llevan a cabo grandes reacciones benéficas plantas-microorganismos en donde las bacterias asociadas al crecimiento de las plantas son denominadas bacterias promotoras del crecimiento en plantas PGPB (Kumar et al., 2016) (Kumar et al., 2017). Los microorganismos antagonistas y sus metabolitos bioactivos representan una de las mejores alternativas a los pesticidas de síntesis química en el control de plagas en cultivos para una agricultura sustentable y seguridad alimentaria (Xu et al., 2019). En general, los microorganismos controladores pueden interactuar con los fitopatógenos mediante diversos mecanismos incluyendo la producción de antibióticos y enzimas líticas, competición por nutrientes, detoxificación de toxinas y degradación de factores de virulencia (Berg et al., 2006). Diversas especies del género Aspergillus son toxigénicas y perjudicial debido a que producen agentes tipo poliquétidos como Aflatoxina B1 (AFB1), B2 (AFB2), G2 (AFG1), G2 (AFG2) y ácido ciclopiazónico (Picot et al.,2017). Los cultivos agrícolas tales como semillas de “algodón”, “maní” y frutos secos son muy propensos a contaminación por aflatoxinas, las cuales son extremadamente carcinogénicas, teratogénicas, mutagénicas y supresoras del sistema inmune cuando son ingeridas incluso en pequeñas cantidades por animales y humanos (Cleveland et al., 2003) (Mobeen et al., 2011). Así también, el oomycete Phytophthora sp. representa a uno de los principales fitopatógenos en los cultivos hortícolas (Kunoba et al., 2016). Entre los candidatos como biocontroladores, las actinobacterias en especial los Streptomyces son de inmenso valor porque ellos son potentes productores de compuestos bioactivos con múltiples propiedades biológicas sumado a sus propiedades filamentosas y de esporulación que les permiten sobrevivir bajo condiciones ambientales desfavorables (Vurukonda et al., 2018). Es por ello la importancia y el objetivo del presente artículo, estudiar actinomicetales aislados de cultivos de “vid” y evaluar su potencial utilidad como PGPB.
Material y métodos
Aislamiento y caracterización enzimática de actinomicetos
Muestras compuestas de suelo fueron colectados de 2 fundos de la ciudad de Ica ubicados según coordenadas: 14°05’00.1”S 75°43’58.5”W, los cuales se caracterizaban por presentar cuadros de “Oidiosis”. Estas muestras fueron procesadas en el laboratorio de Micología Aplicada de UNMSM tomando 10 g de suelo en 90 mL de solución salina, se diluyó hasta 10-1 y se sembró 1 mL por incorporación utilizando como medio el caldo Bushnell Hass (BH) suplementado con carboximetilcelulosa (CMC) al 0,5 % y agar agar 15 g/L. Se realizó por quintuplicado y se incubó a 300C durante 20 días (Evangelista et al., 2014).
Se aislaron las colonias que presentaron las características culturales típicas de actinomicetos (Holt JG et al., 2000) y después de varios pasajes sucesivos en agar BH + CMC se purificaron y se criopreservaron a -200C en agar BH+CMC1% con glicerol al 20%. Para evaluar las características enzimáticas, las cepas fueron evaluadas por su crecimiento en 6 diferentes sustratos tales como: avena, lecitina, caseína, tween 81, almidón y carboximetilcelulosa. Además, se evaluó su crecimiento sobre Agar Nutritivo (AN), Plate count Agar (PCA), PDA (Agar papa dextrosa) y un nuevo medio denominado ASMICO el cual fue elaborado a partir del caldo Bushnell Hass, sacarosa y un aporte de biomasa fúngica dado por el hongo Aspergillus sp.
Cepas fúngicas Testigo
Se utilizaron 2 cepas fúngicas testigo, una de las cuales fue Aspergillus sp. La cual se aisló a partir de muestras de suelo de los cultivos de “vid”, diluidas hasta 10-1 y sembradas en “papa” dextrosa agar (PDA). Se procedió a realizar pasajes sucesivos hasta la purificación de la cepa, la cual finalmente fue identificada como Aspergillus sp. en el Laboratorio de Micología Aplicada UNMSM. La otra cepa fue Phytophthora cinnamomi adquirida del banco de cepas de la Universidad Nacional Agraria La Molina (UNALM).
Preparación de medios de cultivo
Para la preparación del medio ASMICO se utilizó biomasa de la cepa de Aspergillus sp. la cual fue reactivada sembrando una asada en agar PDA, luego de 7 días de incubación se tomaron con sacabocado 5 discos de 8 mm de diámetro y se colocaron en 5 matraces conteniendo caldo Czapeck y sacarosa 15 g/L. Se fermentó durante 7 días a 120 rpm y 30 0C. Los matraces fueron autoclavados a 1210C y 1,5 psi de presión, se tamizó utilizando gasa estéril para separar la biomasa de la fase líquida. Dicha biomasa fúngica obtenida por el proceso fermentativo fue secada en horno a 60 0C durante 3 horas. Finalmente se guardó en frasco estéril hasta su uso. Para preparar el medio de cultivo ASMICO se tomó biomasa seca (1,4 % p/Vol.) 1,4g/100 mL, cloruro de sodio 0,12g/L, cloruro de potasio 0,15g/L, citrato de sodio 0,29 g/L, sacarosa 4,0 g/L y se completó con 15 g/L de agar-agar. Los otros 3 medios de cultivo se elaboraron según su indicación comercial.
Pruebas de antagonismo
En cada uno de los 4 medios de cultivo se colocaron 4 cepas actinomicetales, sembradas por estriado y cada extremo, se incubaron a 300 C durante 72 horas (Zhao et al., 2018). Luego se procedió a colocar la cepa testigo Aspergillus sp. en el centro de la placa. Se sembró también la cepa fúngica testigo sin actinomicetos como control negativo. Todas las placas se sembraron por triplicado y fueron incubadas a 300 C durante 7 días. Luego de este periodo se procedió a verificar cada placa y comparar con el control negativo para verificar la inhibición, el valor de antagonismo se calculó restando el tamaño del diámetro del hongo crecido en el control negativo menos el tamaño del diámetro del mismo Aspergillus sp. que fue enfrentado contra los actinomicetos. A partir de esos resultados se seleccionaron 18 cepas actinomicetales con los mejores resultados de antagonismo. Posteriormente se escogieron las 4 cepas de mayor inhibición y con al menos 1 de las características PGPB y se enfrentaron contra Phytophthora cinnamomi. Estas pruebas fueron realizadas sobre agar PDA siguiendo la misma metodología contra Aspergillus sp.
Evaluación de la capacidad PGPB de actinomicetos
Del total de 35 cepas, se seleccionaron aquellas 18 que presentaron los mejores resultados de antagonismo, estos actinomicetales fueron evaluados para conocer sus características PGPB, tal como se detalla a continuación:
Producción de sideróforos
Las cepas fueron inoculadas en el medio descrito por S & T (Simon & Tessman, 1963), incubadas a 30°C con agitación de 120 rpm por 14 días. Se centrifugó 5 ml del cultivo a 4 000 rpm por 6 min, colocando 100 μL del sobrenadante de cada cultivo con 100 μL de la solución de 5 mM de perclorato férrico (SIGMA-ALDRICH) en 0,14 M de ácido sulfúrico. Para indicar presencia de sideróforos debió ocurrir el cambio de color de la mezcla anterior a un café rojizo.
Fijación de nitrógeno
Las cepas de los actinomicetos fueron sembradas por puntura en Agar NFB, se buscó determinar la capacidad de estos microorganismos para utilizar el N2 como la única fuente nitrogenada, se evaluó así la capacidad de estas actinobacterias para crecer en la superficie de dicho medio (Valdes et al., 2005). Las placas fueron incubadas a 30°C por 14 días, luego de este tiempo se verificó el crecimiento y tamaño de los halos obtenidos.
Solubilización de fosfatos
Las cepas aisladas de actinomicetos fueron reactivadas en caldo bushnell Hass + sacarosa 15%, posteriormente, se sembró cada cepa en placas con medio SRSM-1 (Vásquez et al., 2000) (Gunmarsson et al., 2004), evaluando la presencia de ácidos orgánicos por la variación de pH del medio, lo cual producirá un cambió de color de rojo a amarillo. Las placas fueron incubadas por 14 días a 30°C, luego de este tiempo se midió el tamaño de los halos obtenidos.
Producción de ácido indol acético (AIA)
Los actinomicetos fueron colocados en el Caldo B suplementado con triptona e incubados a 30°C, agitación de 120 rpm durante 14 días. Luego de este tiempo se centrifugó 5 ml de cada cultivo a 3 000 rpm por 15min, se retiró 100 μL del sobrenadante y en un tubo aparte se agregó 200 μL del reactivo de Salkowsky, finalmente, se dejó en oscuridad durante 15 minutos. La lectura positiva para indicar presencia de AIA fue dada por el viraje de color, de amarillo a fucsia (Celis et al., 2007).
Análisis estadístico
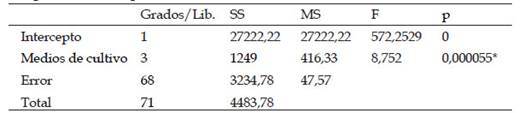
Mediante el análisis de varianza (ANOVA) utilizando el software Statistica 10, se comparó la influencia de los diferentes medios de cultivo en la producción de metabolitos antifúngicos verificando así si existe diferencias significativas entre los diferentes medios de cultivo. Para ello se estableció la siguiente hipótesis: H0: No hay diferencia significativa entre los grupos evaluados (µ1= µ2= µ3= µ4) Es decir los medios de cultivo no influyen en la respuesta antagonista de los actinomicetos, H1: Al menos uno de los grupos evaluados es significativamente diferente (µ1≠ µ2≠ µ3≠ µ4). Es decir, los medios de cultivo sí influyen en la respuesta antagonista de los actinomicetos. Se utilizó un alfa α=0.05
Resultados
Aislamiento y caracterización enzimática de actinomicetos
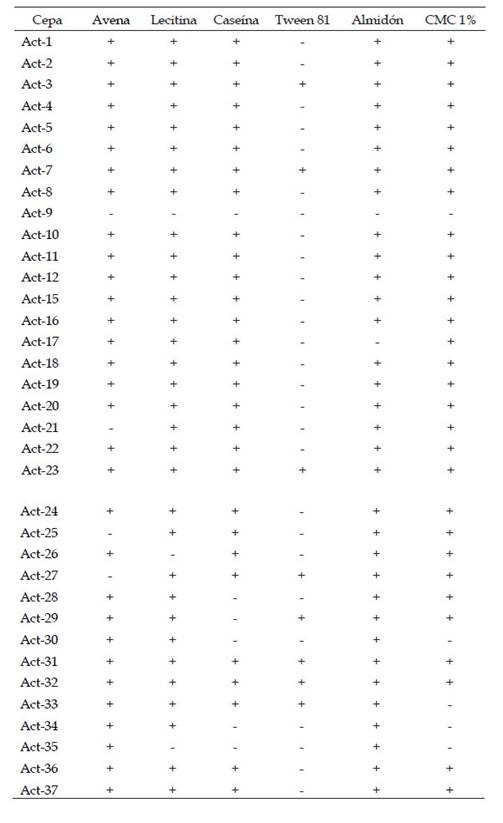
35 actinomicetos fueron aislados de suelos de cultivos de “vid” Vitis vinifera (Vitaceae) en Ica, Perú. Debido a sus características culturales típicas en los diversos medios de cultivo, entre ellos el agar con CMC, que les permiten un crecimiento bien diferenciado con otros grupos microbianos, se pudo lograr un aislamiento sin demasiadas purificaciones, favoreciendo así un aislamiento más eficiente. En resumen, este estudio permitió confirmar que 31 cepas (88,6%) fueron capaces de degradar avena, 32 cepas (91,4%) poseen lecitinasas, 29 cepas (82,9%) degradan caseína, 8 cepas (22,9%) poseen esterasas, 33 (94,3%) cepas poseen amilasas y 30 cepas (85,7%) degradan CMC. (Tabla 1)
Influencia de los medios de cultivo
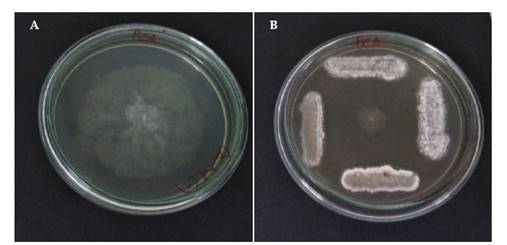
Los 4 medios de cultivo evaluados permitieron evidenciar que el poder antagonista de los actinomicetos varía dependiendo del medio en el cual se desarrolla la prueba. Así se pudo comprobar que el medio ASMICO, elaborado con biomasa de Aspergillus sp., sales y sacarosa fue el que presentó los mejores resultados en comparación a PDA, PCA y AN. En la Figura 1 se puede apreciar el poder antagonista de algunos de los actinomicetos seleccionados, en los diferentes medios de cultivo. Se confirmó que en los 4 medios de cultivo existe inhibición de Aspergillus sp. sin embargo fue el medio de cultivo ASMICO el que permitió obtener los mejores resultados de antagonismo.
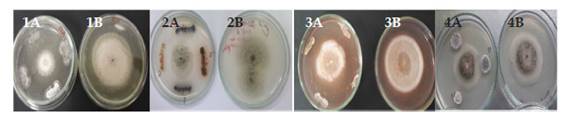
Pruebas de antagonismo
De las 35 cepas de actinomicetos aisladas, 18 fueron seleccionadas debido a que presentaron inhibición contra la cepa testigo Aspergillus sp. (Tabla 2). La respuesta antagonista en los 4 medios de cultivo utilizados se evidencia con la Figura 1. El medio de cultivo ASMICO fue el que presentó las medias más altas con 26,2 mm como tamaño promedio por la inhibición en el crecimiento del hongo testigo (Figura 2). En base a los mayores tamaños de inhibición provocado contra Aspergillus sp. 4 cepas de actinomicetos fueron seleccionadas Act-1, Act-11, Act-17 y Act-22. Estos 4 actinomicetos provocaron la inhibición de Phytophthora cinnamomi, reduciendo su crecimiento hasta 10 mm de diámetro en comparación a los 49 mm del control negativo, reduciendo en 39 mm el crecimiento del fitopatógeno testigo (Figura 3).
Tabla 2 Tamaños (mm) obtenidos por la inhibición del crecimiento de Aspergillus sp.
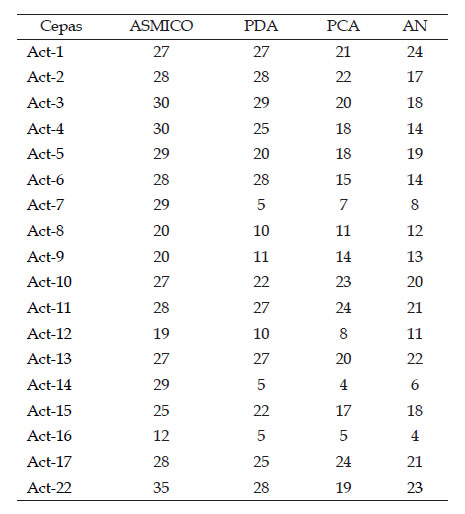
*Los valores son el promedio de 3 repeticiones.
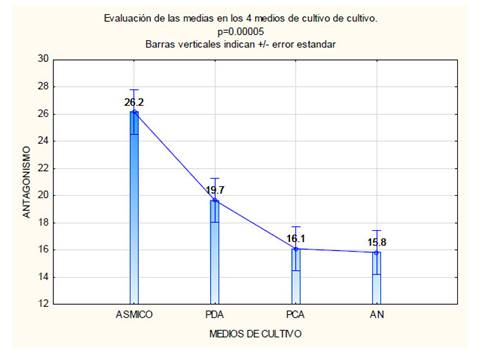
Fig. 2 Comparación de las medias de los antagonismos (parte superior de las barras) obtenidas en cada uno de los medios de cultivo. ASMICO se diferencia de los otros 3 medios comerciales.
Evaluación de la capacidad PGPB de actinomicetos
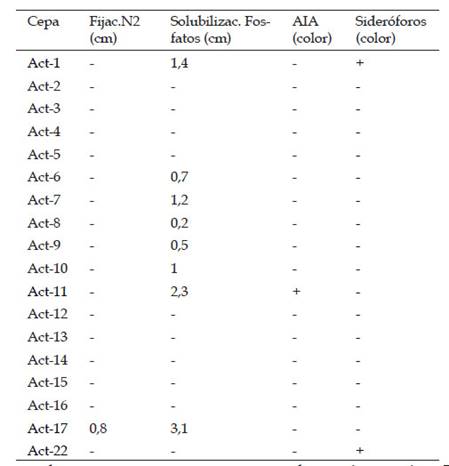
Se confirmó que 8 actinomicetos presentaron capacidad para solubilizar fosfatos. 2 actinomicetos con capacidad para producir sideróforos fueron seleccionados, las cepas Act-1 y Act-22, y debido a que también presentaron poder antagonista, fueron seleccionadas. La cepa Act-11 fue seleccionada debido a la producción de AIA y por ser una cepa con poder antagonista. Y finalmente la cepa Act-17 presentó propiedades para fijar nitrógeno, por ello fue la última en ser seleccionada. Con estos resultados se seleccionaron las 4 cepas que presentaron tanto actividad antifúngica, así como al menos 1 de las propiedades PGPB. En la tabla 3 se resumen las características PGPB de las 18 cepas seleccionadas.
Análisis estadístico
Mediante al análisis ANOVA se confirmó la existencia de diferencias significativas entre los 4 medios evaluados ya que se obtuvo un p <0.05 (Tabla 3), por lo cual se rechaza la hipótesis nula y se acepta la hipótesis alternativa. El análisis Tukey evidenció que el medio ASMICO es el único que muestra dicha diferencia (Tabla 4).
Tabla 4 Prueba de Tukey HSD para la variable Antagonismo. Df=68, MS=47.57
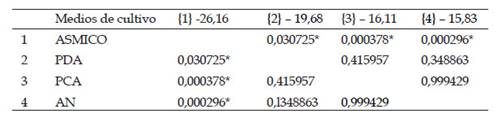
Indican que la respuesta fue significativa, p<0,05
Discusión
Debido a las características culturales típicas de los actinomicetos en los diversos medios de cultivo, entre ellos el agar con CMC, que les permiten un crecimiento bien diferenciado con otros grupos microbianos, se pudo lograr un aislamiento sin demasiadas purificaciones, favoreciendo así un proceso más eficiente. Dichos aislamientos fueron favorecidos por la utilización de carboximetilcelulosa (CMC) al 0,5% ya que a pesar de ser una fuente carbonada que limita el crecimiento de gran parte de flora acompañante, sí es asimilable por los actinomicetos (Grigorevski et al.,2005). Mientras que el nitrato de amonio y las demás sales (Sulfato de magnesio, cloruro de calcio, fosfato monopotásico, fosfato dipotásico y cloruro férrico) aportadas por el medio Bushnell Hass (BH) actuaron como fuente nitrogenada y suplemento de micronutrientes, respectivamente. Al utilizar un medio con baja concentración de nutrientes y una fuente carbonada de difícil degradación por la mayoría de flora bacteriana se pudo utilizar la dilución más concentrada 10-1 mejorando así los tiempos de aislamiento y purificación de las cepas. A diferencia de autores como Gansean et al. (2017). en donde realizó diluciones seriadas hasta 10-4 ó 10-5 además de utilizar medios altamente enriquecidos con fuentes carbonadas y nitrogenadas e incluso antifúngicos. La tabla 1 revela una gran capacidad por parte de los actinomicetos para excretar enzimas líticas. Estos resultados permiten confirmar una elevada expresión y secreción de enzimas lo cual favorece una mejor respuesta antagonista ya que muchas veces estos metabolitos pueden interactuar directamente sobre fitopatógenos causando su inhibición (Pal et al., 2006). Además, esta característica multienzimática les permite degradar mayor cantidad de sustratos presentes en el suelo y por lo tanto sobrevivir y adaptarse mejor.
Cuando se comparó la influencia de los 4 medios de cultivo sobre la respuesta antifúngica contra Aspergillus sp. se observó con claridad que el nuevo medio de cultivo ASMICO favorece la producción de antifúngicos a pesar que las colonias de los actinomicetos fueron más pequeñas en este medio en comparación a los otros 3. Esto indica claramente que la producción de biomasa microbiana no necesariamente nos indica una mayor producción de antifúngicos. Esto se puede explicar porque el metabolismo microbiano en un medio con alta cantidad de nutrientes se direcciona hacía el crecimiento sin embargo en un medio con baja cantidad de nutrientes se activan rutas metabólicas para la producción de metabolitos secundarios con propiedades antifúngicas. Esto explicaría el mayor efecto antagonista en el nuevo medio de cultivo. Esta inhibición mostró diferentes comportamientos del hongo testigo ya que, aunque pudo crecer con facilidad en los 4 medios de cultivo estudiados fue en los medios ASMICO y PDA en donde el tamaño de sus colonias fue reducido con mayor notoriedad, minimizando incluso la altura del micelio aéreo. Estos resultados son congruentes con lo demostrado por Grahovac et al. (2014) quienes afirmaron que la producción de antifúngicos varía dependiendo de los nutrientes que se utilizan para el crecimiento. Mediante la prueba de HSD-Tukey se confirmó que de los 4 medios de cultivo evaluados fue el medio ASMICO el único que presentó diferencia significativa (Tabla 4), con lo cual se afirma que este nuevo medio de cultivo es un inductor en la producción de antifúngicos en mayor medida que los otros 3 medios comerciales utilizados. Lo cual también está directamente relacionado con el stress nutricional inducido por la poca cantidad de nutrientes que a su vez provocan la activación de rutas metabólicas que desencadenen en la producción de antimicrobianos (Lino et al., 2016). Debido a la limitación de nutrientes tanto en variedad como en concentración en el medio ASMICO se favoreció el poder antagonista de la mayoría de cepas actinomicetales evaluadas. Las 4 cepas actinomicetales seleccionadas nos permitieron afirmar que dichas cepas producen compuestos antifúngicos capaces de inhibir también el crecimiento de Phytophthora cinnamomi. Este poder antagonista se hizo evidente con los 39 mm de reducción en el crecimiento de dicho fitopatógeno. El medio ASMICO fue el más idóneo para realizar dicha selección ya que permitió encontrar una mayor cantidad de cepas actinomicetales con potencial para excretar metabolitos antifúngicos. Ya que si no se hubiese realizado las pruebas de antagonismo en este medio la cantidad de cepas seleccionadas hubiese sido menor.
El 44,4% de actinomicetos presentaron capacidad para solubilizar fosfatos, y fue esta la mejor respuesta de todas las 4 características PGPB evaluadas. Dentro de estas 8 actinobacterias, se encontraron 3 cepas que presentaron los mejores resultados de antagonismo, esto permitió seleccionarlas debido a que son productores de antifúngicos y son capaces de solubilizar fosfatos. Dentro de los microorganismos solubilizadores de fosfatos también se destacan Acinetobacter sp. Bacillus sp. y Pseudomonas sp. (Joe et al., 2016). Estos microorganismos, así como un gran número de actinobacterias son consideradas promotoras del crecimiento en plantas por lo que es importante su estudio ya que permiten mejorar la producción, así como reducir el consumo de fertilizantes. La evaluación de sideróforos es un factor importante, ya que facilitan el ingreso de hierro a las plantas. Al evaluar la producción de ácido indol acético AIA, nos permitió seleccionar cepas productoras de esta fitohormona que está directamente involucrada en la diferenciación celular, la división celular, así como en la elongación de las plantas (Goswami et al., 2016). Nuestros resultados confirmaron que la cepa Act-11 es un actinomiceto productor de AIA, por ello y por ser una cepa con poder antagonista esta cepa también fue seleccionada. El Nitrógeno está presente en el suelo de forma abundante, pero en forma no asimilable para las plantas, ya que las plantas lo consumen bajo la forma de amonio (NH4) y nitrato (NO3). La conversión de nitrógeno atmosférico N2 hasta dichas formas asimilables tales como amonio (NH4) y nitrato (NO3) es conocido como la fijación de nitrógeno, dicha habilidad está presente en bacterias y arqueas (Das et al., 2015), nuestros resultados permitieron aislar y seleccionar a la cepa Act-17 debido a que se le encontró está característica y también produce metabolitos antifúngicos. Debido a estos resultados los actinomicetos seleccionados fueron 4 y codificados como Act-1, Act-11, Act-17 y Act-22. Por lo anterior se afirma que es muy importante evaluar diferentes medios de cultivo para una buena selección de cepas actinomicetales productoras de antifúngicos que inhiben tanto a Aspergillus sp. así como a Phytophthora cinnamomi así como para seleccionarlas como potenciales bacterias promotoras de crecimiento en plantas o también llamadas PGPB. Finalmente, estos resultados nos brindan una técnica sencilla para aislar actinomicetos de cultivos de “vid” los cuales pueden ser utilizados tanto en el biocontrol de fitopatógenos así como estimulantes del crecimiento en plantas, en pro de una agricultura sustentable y con menos agroquímicos.
Conclusiones
Los actinomicetos aislados de cultivos de “vid” Vitis vinifera (Vitaceae) en Ica, producen metabolitos antifúngicos capaces de inhibir a Aspergillus sp. y Phytophthora cinnamomi. Además, estas actinobacterias poseen características que les confiere pertenecer al grupo de bacterias promotoras de crecimiento de plantas PGPB. Es importante utilizar diferentes medios de cultivo para evaluar el poder antagonista de las cepas actinomicetales ya que dicha inhibición puede verse aumentada o disminuida por el tipo y concentración de los nutrientes que se utilicen.

