











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Los bosques montanos de la vertiente oriental de los Andes presentan un mosaico de tipos de vegetación, que responden a diferencias en la precipitación, estacionalidad, latitud y altitud (K. Young et al., 2007). La interacción de estos factores, junto con la presencia de neblina, fuertes pendientes y variados regímenes de temperatura condicionan la existencia de una variedad de microclimas que favorecen la alta diversidad biológica en estos ecosistemas (Young & León, 1995). La vertiente oriental de los Andes peruanos ha sido reconocida a nivel global por su alta biodiversidad (Brummitt & Lughadha, 2003) y contiene numerosas zonas ecológicas distribuidas a lo largo de 5000 metros de elevación en una zona muy estrecha que va de norte a sur a lo largo de diez grados en los trópicos (Young & León, 1995). El bosque montano alto se ubica principalmente en los Andes orientales entre los 2500 m hasta el límite del bosque que varía entre los 3600 a 3800 m de elevación (Ministerio del Ambiente, 2018). A medida que se incrementa la elevación, la altura del dosel del bosque disminuye, los árboles son más torcidos y nudosos, tienden a tener tallos múltiples (Young et al., 2007), se incrementa el área basal, pero disminuye la riqueza de especies (Malizia et al., 2020).
De manera general, estos bosques se caracterizan por la presencia de familias como Lauraceae, Melastomataceae, Myrtaceae, Rubiaceae, y Solanaceae en zonas más húmedas y de Fabaceae y Asteraceae en los bosques con menos humedad (Young et al., 2007). Las plantas epífitas como Orchidaceae y Bromeliaceae, así como musgos y líquenes son abundantes. En el Perú, se ha identificado a Podocarpus (Podocarpaceae), Hedyosmum (Chloranthaceae) y Weinmannia (Cunoniaceae) como géneros dominantes de árboles, junto con una alta abundancia de helechos arbóreos (Reynel et al., 2013).
En años recientes ha habido esfuerzos para caracterizar estos bosques a niveles continentales, a través de redes de colaboración (Báez et al., 2015; Girardin et al., 2014) y en el Perú hay esfuerzos locales que están incrementando nuestro conocimiento en términos de la biodiversidad y ecología de estos bosques usando redes de parcelas a lo largo de gradientes de elevación (Antón & Reynel, 2009; Bañares de Dios et al., 2020; Farfán-Rios et al., 2015; Valenzuela et al., 2015). Sin embargo, dado que estos bosques cubren una superficie de casi 24000 km2 (Ministerio del Ambiente, 2018) y se extienden por más de 1500 km desde Piura, Amazonas y Cajamarca en el norte hasta Puno en el sur, aun quedan espacios geográficos de los cuáles no conocemos adecuadamente su diversidad florística y menos su estructura boscosa (Honorio & Reynel, 2012). En ese sentido, este trabajo tiene como objetivo principal contribuir con datos de inventario cuantitativos de especies leñosas en parcelas con un área y metodología estándar en los bosques altimontanos de yunga del suroeste del Parque Nacional Rio Abiseo.
Materiales y métodos
Área de estudio
El área de estudio se localiza al norte del Perú en el distrito de Huicungo, provincia de Mariscal Cáceres, región San Martín. Geográficamente se ubica en el sector occidental del Parque Nacional Río Abiseo (PNRA) en la vertiente oriental de la cordillera de los Andes, en la zona denominada La Sata a una altitud promedio de 2700 msnm. El Mapa Nacional de Ecosistemas (Ministerio del Ambiente, 2018), define al área de estudio como Bosque Altimontano de Yunga (Figura 1). La zonificación del PNRA correspondiente es de Zona Histórico-Cultural (Figura 1).
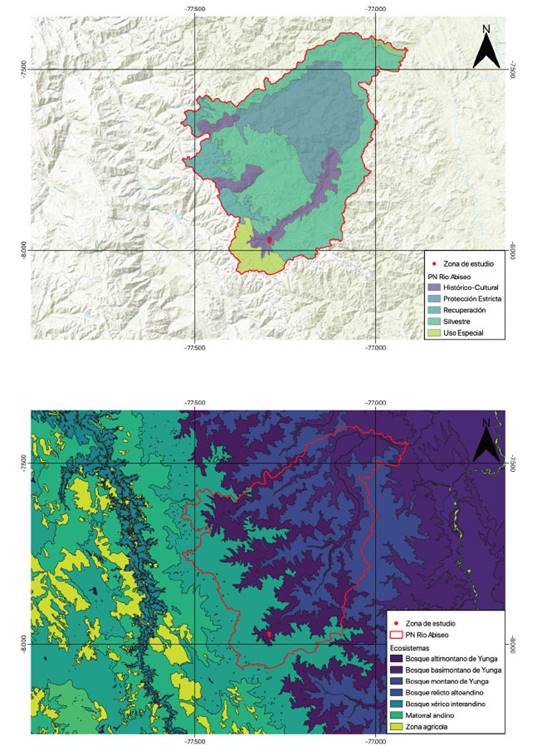

Fig. 2 Tipos de vegetación evaluados. De arriba abajo, y de izquierda a derecha: Pajonal, bosque con sotobosque apisonado, bosque de ladera y bosque remanente.
Hay dos estaciones bien marcadas una de octubre a abril, cuando ocurren las mayores precipitaciones y la otra entre mayo y septiembre, cuando la temperatura desciende y la precipitación disminuye. La precipitación promedio anual para la cuenca del río Montecristo, zona cercana al área de estudio, probablemente sea de 2500 mm (Young, 1993).
Muestreo de la vegetación
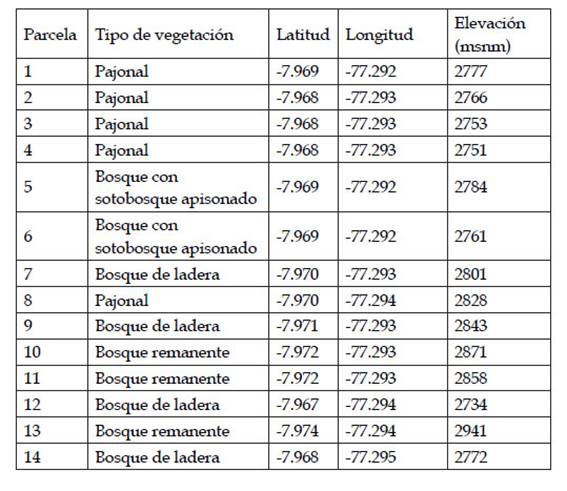
El 2018 se instalaron 14 parcelas modificadas Whittaker de 0,1 ha (Stohlgren et al., 1995) en dos unidades y cuatro zonas, que fueron re-visitadas el 2019 (Tabla 1). Las unidades son bosque intervenido y bosque no intervenido. La unidad de bosque intervenido se dividió en tres zonas: 1) Bosque de ladera donde hubo extracción selectivademaderaprevioalestablecimiento del PNRA, 2) Bosque con sotobosque apisonado por actividad ganadera y extracción maderera selectiva previa al establecimiento del PNRA, y 3) Pajonal, usado como zona de pastoreo y agricultura extensiva previo al establecimiento del PNRA. La unidad de bosque no intervenido es la zona con vegetación remanente o zona donde identificamos la menor intervención antrópica posible previo al establecimiento del PNRA.
Cada parcela Whittaker modificada mide 20 m x 50 m (Parcela C, Figura 3). Estas fueron instaladas siguiendo el manual de instalación de parcelas permanentes de Phillips et al. (2016). El Vértice 1 de cada parcela se ubica en la esquina inferior derecha, tomando en cuenta la dirección de la pendiente. Cada parcela a su vez fue dividida en dos tipos de subparcelas anidadas, cada una de tamaño y ubicación diferente. El primer tipo (Parcela A) contiene 10 subparcelas separadas (de 0.5m x 2m cada una). El segundo tipo (Parcela B) incluye una sola subparcela en la parte central (de 5 m x 20 m).
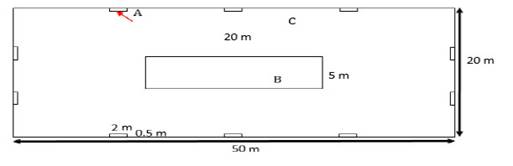
En cada Parcela A se estimó la cobertura relativa (en porcentaje) de todas las especies herbáceas, donde un área de 10cm x 10cm equivale al 1% de cobertura. En la parcela B se registró todas las especies arbustivas con 1cm ≥ DAP < 5cm. En la parcela C se registró todas las especies presentes con un DAP ≥ 5cm y sus respectivas alturas por estimación visual. Para la medición del DAP se limpió la corteza de los árboles y la medida fue tomada a 1.3m con una cinta diamétrica. En el caso de árboles con fuste irregular, es decir, con abultamientos o tallo acanalado, la medición del DAP se hizo en la superficie más homogénea posible por encima de 1,3 m. Estos detalles fueron anotados en el cuaderno de campo.
Se colectaron muestras de plantas en los meses de julio y agosto del 2018 y 2019. Se etiquetó cada muestra con el número correspondiente al árbol colectado. Las muestras fueron preservadas en alcohol al 70% hasta su secado en el Herbario Vargas de la Universidad Nacional de San Antonio Abad del Cusco (UNSAAC, Cusco). La identificación de las muestras de plantas se realizó con ayuda de especialistas, visita a herbarios (USM, CUZ). Se realizaron comparaciones con ejemplares de los herbarios virtuales de Missouri Botanical Garden (https://www.tropicos.org), del Field Museum of Natural History (https://collections-botany.fieldmuseum. org), JSTOR Global Plants (https://plants. jstor.org) y ATRIUM (http://atrium. andesamazon.org/digital_herbarium. php). También se revisó bibliografía especializada. La nomenclatura sigue a APG IV (The Angiosperm Phylogeny Group et al., 2016).
Análisis de datos
Se compiló las especies registradas en cada tipo de vegetación en base a los datos de las especies leñosas con un DAP ³ 5 cm. Se calculó el número de árboles, el área basal y la distribución del número de árboles por clases diamétricas y de altura. Para estos dos últimos, también se incluyó la información de los individuos con DAP ³ 1cm. Calculamos el Índice de valor de Importancia (IVI), para medir el “valor ecológico” de cada especie en cada tipo de vegetación sumando los valores relativos de la dominancia (área basal), la frecuencia (ocurrencia de cada especie en las diferentes parcelas del mismo tipo de bosque) y la abundancia de cada especie (Curtis & McIntosh, 1951).
Resultados y discusión
Composición florística, riqueza de especies y estructura del área de estudio
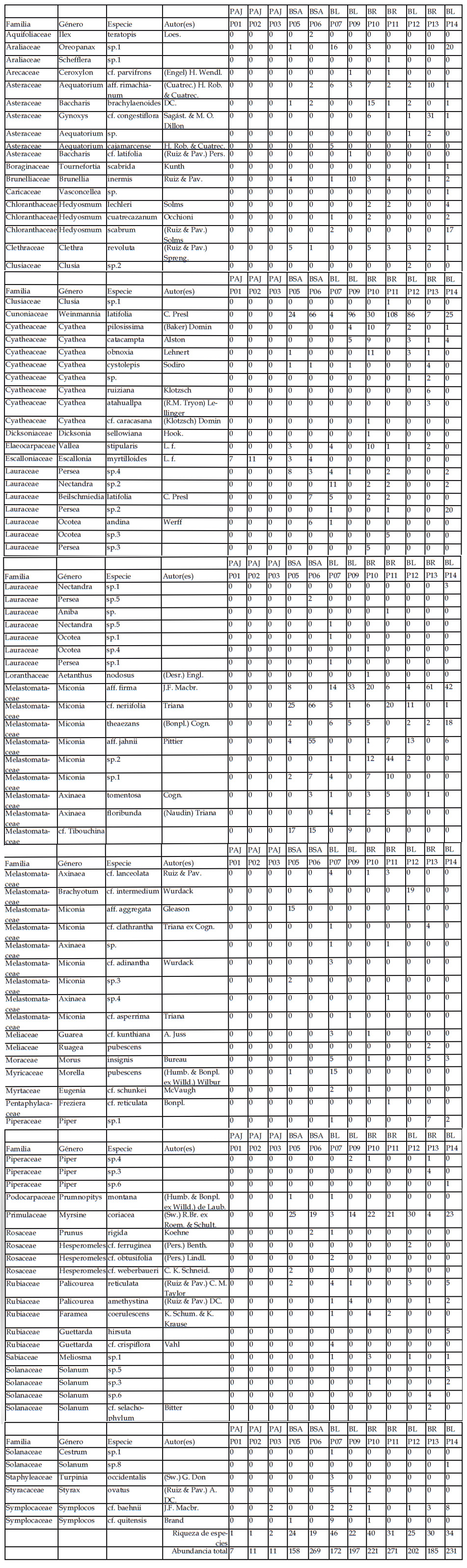
Registramos 96 especies leñosas en las 12 de las 14 parcelas estudiadas (en 33 familias y 48 géneros), incluyendo morfoespecies (Tabla 1). Dos parcelas en pajonales no contuvieron especies leñosas (P04, y P08). La familia más diversa fue Melastomataceae (18 especies), seguida de Lauraceae (14) y Cyatheaceae (7). Las parcelas con mayor número de especies fueron las parcelas P07 y P10 (45 y 40 especies, en bosques de ladera y remanentes, respectivamente), las parcelas con menos especies fueron las parcelas en pajonales (1-3 especies). Los valores de densidad (número de individuos) fueron más altos en las parcelas P11 y P06 (271 y 269 individuos, respectivamente). En la primera dominó Weinmannia latifolia C. Presl (108 individuos), seguida de Miconia sp.2 (44 ind.). en la segunda parcela dominaron Weinmannia latifolia C. Presl (66 ind.), Miconia cf. neriifolia Triana (66 ind.) y Miconia aff. jahnii Pittier (55 ind.). De manera general, Weinmannia latifolia C. Presl fue la especie más abundante en la zona de estudio (446 individuos), seguida de Miconia aff. firma J. F. Macbr. (188 ind.), Myrsine coriacea (Sw.) R. Br. ex Roem. & Schult. (161 ind.) y Miconia cf. neriifolia Triana (135 ind.). Conjuntamente, estas cuatro especies contribuyeron al 48% del total de individuos. Por el contrario, registramos 18 especies con sólo un individuo. Las especies que ocurrieron en más parcelas fueron Weinmannia latifolia C. Presl y Myrsine coriacea (Sw.) R.Br. ex Roem.
& Schult. (ambas presentes en 9 parcelas), seguidas de Aequatorium aff. rimachianum (Cuatrec.) H. Rob. & Cuatrec., Brunellia inermis Ruiz & Pav., Miconia aff. firma J.F. Macbr. y Miconia cf. neriifolia Triana (todas con presencia en 8 parcelas).
Considerando cada uno de los tipos de vegetación estudiados, en la zona del pajonal, sólo registramos tres especies leñosas, dominando Escallonia myrtilloides (Tabla 2). En el bosque con sotobosque apisonado registramos 32 especies (en 24 géneros y 18 familias). De las especies colectadas, el 50% fueron asignadas a morfoespecies. Las familias con mayor número de individuos fueron Cunoniaceae, Melastomataceae, Primulaceae y Lauraceae. Registramos 448 individuos, siete de los cuales se encuentran como indeterminados. Las familias representadas por un solo género son: Aquifoliaceae, Araliaceae, Brunelliaceae, Elaeocarpaceae, Clethraceae, Escalloniaceae, Myricaceae, Podocarpaceae, Primulaceae, Rubiaceae y Symplocaceae. El género Miconia presentó el mayor número de especies (7) (Tabla 2). En el bosque de ladera registramos 78 especies (en 40 géneros y 29 familias), 61,5% de las cuáles fueron reconocidas como morfoespecies. De los 797 individuos registrados, ocho no pudieron ser identificados. Las familias con mayor número de especies fueron Asteraceae, Chloranthaceae, Cyatheaceae, Lauraceae, Melastomataceae, Rubiaceae, Solanaceae, Piperaceae y Rosaceae. El género con mayor número de especies fue Miconia (11), seguido por Axinaea (5), Cyathea (5), seguido por Hedyosmum, Persea, Nectandra y Solanum cada uno con tres especies (Tabla 2). En el bosque remanente registramos 67 especies (en 36 géneros y 26 familias), con un 59,7% de morfoespecies. Del total de individuos registrados (685), ocho individuos se encuentran sin determinar. Las familias con mayor número de géneros fueron Asteraceae (3), Lauraceae (5), seguidas por Melastomataceae, Rubiaceae, Meliaceae y Araliaceae, cada una con dos géneros respectivamente. Los géneros con mayor número de especies fueron Cyathea (7), Miconia (7), Persea (3), Axinaea (5), Solanum (4) y Piper (3) (Tabla 2).
Tabla 2 Abundancia de especies leñosas en parcelas de La Sata, Parque Nacional Rio Abiseo. PAJ = Pajonal, BSA = bosque con sotobosque
La distribución de la abundancia muestra que el bosque de ladera y el bosque remanente tienen una mayor cantidad de individuos en categorías diamétricas intermedias (10-29,9cm) y por lo tanto mayor área basal que los otros tipos de bosques (Fig. 4). En el pajonal se observó mayor número de individuos con diámetro menor igual 4,9 cm que el en bosque remanente. Sin embargo, estas parcelas tuvieron una menor abundancia en la clase diamétrica 5-9,9 cm, esto debido probablemente a factores limitantes en la dispersión de semillas y en la germinación. Considerando sólo las clases diamétricas de 5 cm de DAP y superiores, se observó bosques con individuos de edades disparejas reflejadas en la clásica curva con forma de J invertida, con mayor cantidad de individuos en clases diamétricas menores y muy pocos individuos en clases diamétricas mayores. Esto sugiere que los bosques de La Sata son viables y que hay adecuada regeneración (Newton, 2007).
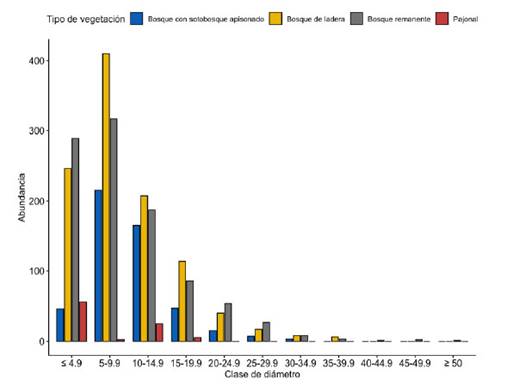
Fig. 4 Distribución de número de individuos en clases de diámetro. Los individuos con diámetro menor a 5cm sólo se muestrearon en la subparcela de 5 m x 20 m.
La distribución de los individuos por categorías de altura en cada tipo de vegetación mostró que en el pajonal todas las leñosas registradas tienen alturas menores o iguales a 9,9 m. En el bosque de ladera la mayor parte de los individuos tuvieron alturas entre 5 y 14,9 m. En el bosque remanente los individuos se concentraron en clases de altura menores a 4,9 y 10-14,9 m, y es el que también tuvo un mayor número de individuos con alturas iguales o mayores a 15m (Fig. 5).
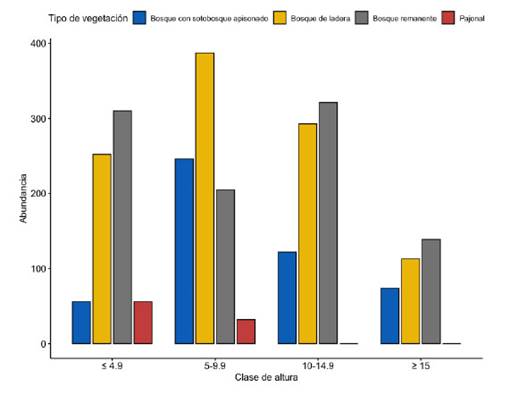
En el pajonal, la especie que tuvo mayor índice de valor de importancia (IVI) fue Escallonia myrtilloides con 241,02% (Tabla 3). Esto valor alto de IVI resultado de valores relativos altos de abundancia y de área basal (ambos 90% o superior). En la zona denominada bosque con sotobosque apisonado, las especies con mayor índice de valor de importancia fueron Weinmannia latifolia (49,23%) y Miconia cf. neriifolia (41,77%), seguidas por Miconia aff. jahnii (26,25%) y Myrsine coriacea (24,20%) (Tabla 4). Las dos primeras tuvieron valores altos de números de individuos (90 y 91 individuos, respectivamente), y el área basal de W. latifolia incluyó casi un cuarto del total para medido este bosque (1,23 m2). En el bosque de ladera la especie dominante en base al valor de IVI fue Weinmannia latifolia (58,81). Le siguen Miconia aff. firma (28,68%) y Myrsine coriacea (16,31%) (Tabla 5). W. latifolia mostró valores altos de abundancia (211 individuos) y área basal (3,04m2), constituyendo en ambos casos entre el 26 y 29% de los valores totales para este tipo de bosque. Finalmente, en el bosque remanente Weinmannia latifolia y Miconia aff. firma dominaron con los valores más altos de IVI (38,09% y 35,65%), seguidas de Gynoxis cf. congestiflora (18,32%), Miconia sp. 2 (15,14%) y Myrsine coriacea (12,70%.) (Tabla 6). W. latifolia dominó los valores de abundancia (144 individuos, 21% del total), mientras que Miconia aff. firma dominó los valores de área basal (2,14m2, 20% del valor total) en este tipo de bosque.
Tabla 3 Índice de Valor de Importancia (IVI) de especies leñosas en el pajonal (5 parcelas). A = Abundancia de individuos, D = Dominancia expresada por el área basal (m2), F = frecuencia, AR = Abundancia Relativa, DR = Dominancia Relativa, FR = Frecuencia relativa.
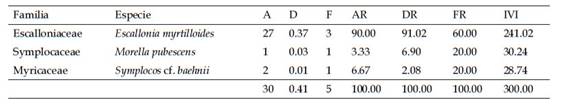
Tabla 4 Índice de Valor de Importancia (IVI) de las diez especies leñosas con valores más altos en el bosque con sotobosque apisonado (2 parcelas). A = Abundancia de individuos, D = Dominancia expresada por el área basal (m2), F = frecuencia, AR = Abundancia Relativa, DR = Dominancia Relativa, FR = Frecuencia relativa.
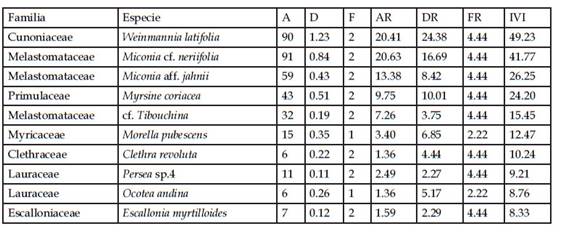
Tabla 5 Índice de Valor de Importancia (IVI) de las diez especies leñosas con valores más altos en el bosque de ladera (4 parcelas). A = Abundancia de individuos, D = Dominancia expresada por el área basal (m2), F = frecuencia, AR = Abundancia Relativa, DR = Dominancia Relativa, FR = Frecuencia relativa.
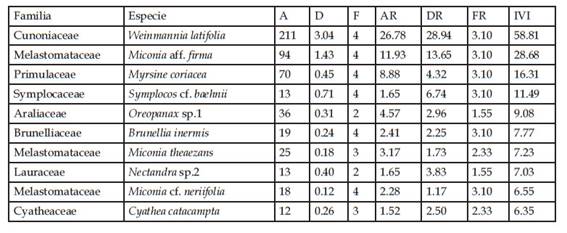
Tabla 6 Índice de Valor de Importancia (IVI) de las diez especies leñosas con valores más altos en el bosque remanente (3 parcelas). A = Abundancia de individuos, D = Dominancia expresada por el área basal (m2), F = frecuencia, AR = Abundancia Relativa, DR = Dominancia Relativa, FR = Frecuencia relativa.
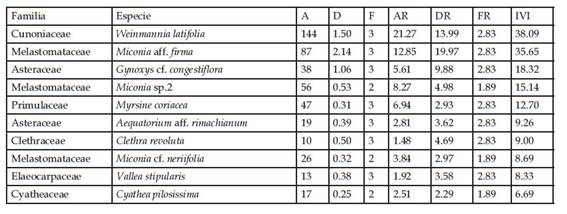
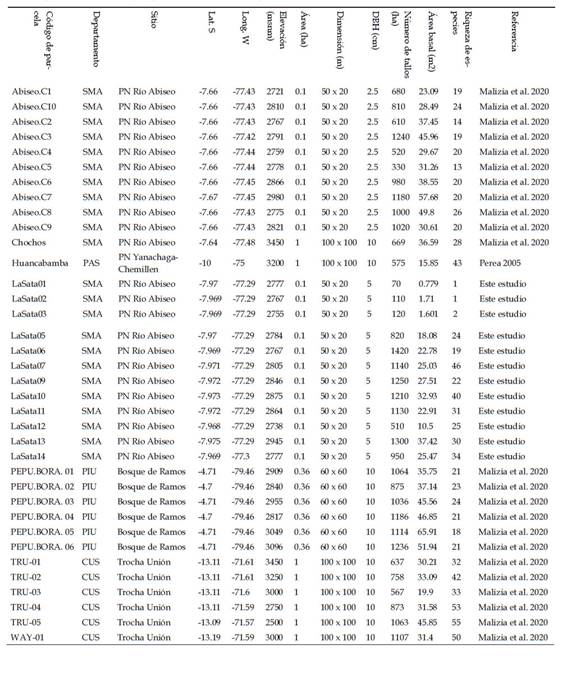
En comparación con otros inventarios de bosque altimontano de yungas por encima de los 2500 m de elevación en el Perú (Tabla 7), los bosques de La Sata tendrían los valores más altos de densidad de tallos con DAP >= 10 cm. Las parcelas P06, P13 y P09 todas presentaron valores por encima de 1250 tallos por hectárea. De la misma manera, tres parcelas adicionales (P07, P10 y P11) registraron valores superiores a 1100 tallos por hectárea, con lo cual seis de las 14 parcelas evaluadas en este estudio presentan los valores de densidad más altos hasta ahora registrados en bosques montanos. Es interesante notar que, en este grupo de parcelas con valores de densidades altos, están también dos parcelas adicionales inventariadas en el PNRA. Sin embargo, ninguna de las parcelas evaluadas en este estudio presentó valores altos de área basal, excepto la parcela P13 con un valor medianamente alto (37.42m2). Finalmente, en términos de riqueza de especies, dos parcelas de este estudio obtuvieron valores mayores o iguales a 40 especies (en 0,1 ha).
Tabla 7 Parcelas de inventarios de árboles en bosques altimontanos de yungas en el Perú por encima de los 2500 msnm. El número de tallos está extrapolado a una hectárea para parcelas de menor tamaño, y sólo considerando tallos con DAP ≥ 10cm.
La comparación de los resultados florísticos, muestra que las parcelas de La Sata coinciden de manera general con los patrones de dominancia de árboles en otros bosques del mismo ecosistema en el Perú. Melastomataceae, Lauraceae y Cyatheaceae (incluida en Pteridophyta en algunos estudios) son las familias más especiosas registradas tanto en nuestro estudio, como en los bosques de Junín (de Rutte & Reynel, 2016; Reynel & Antón, 2004; Reynel & Honorio, 2004), Cusco (Rivera, 2007), San Martín (Young, 1998) y Pasco (Gómez, 2000; Perea, 2005). De igual manera, nuestros datos coinciden con estos estudios en que Miconia y Weinmannia son los géneros más abundantes.
Conclusiones
Los bosques montanos son una prioridad global de conservación debido a su gran diversidad y alto nivel de endemismo (Pennington et al., 2010) a su vez brindan servicios ecosistémicos como la regulación del clima, protección de especies, participan en el ciclo hidrológico y de carbono; a pesar de ello, este ecosistema es uno de los más amenazados (Kessler, 2000; Mathez-Stiefel et al., 2017). Considerando que aún tenemos deficiencias en nuestro conocimiento de qué especies existen en estos bosques, el llamado Déficit Linneano, y de su distribución, el Déficit Wallaceano (Brito, 2010), se debe seguir trabajando en describir las características básicas de composición y estructura de estos bosques. Acompañado de incrementos en las colectas botánicas, mejorará los niveles de determinación de las especies. A pesar de que realizamos el esfuerzo de visitar el sitio de estudio durante dos años consecutivos, aun tuvimos muchas especies que solo pudieron ser colectadas como especímenes infértiles. Es claramente necesario invertir esfuerzos en parcelas de muestreo permanente que permitan monitorear la dinámica del bosque en su respuesta frente a cambios globales (Feeley et al., 2020).

