Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Carica papaya (Caricaceae) “papaya”, es un frutal originario del Noroccidente Amazónico y distribuida actualmente por todos los trópicos y subtrópicos a nivel mundial. Es una planta de crecimiento relativamente rápido y de vida corta cuyo cultivo dura de 1 a 3 años (Mostacero et al., 2002). Esta planta puede ser cultivada desde el nivel del mar hasta los 1000 msnm, dependiendo de las condiciones climáticas. Soportando entre 22° y 30°C, pero su óptima es entre 23° y 26°C (Rodríguez, 2004). Su cultivo se ha incrementado considerablemente y su demanda ha aumentado especialmente en mercados internacionales en años recientes, ya sea en su forma fresca o como producto procesado (Sankat & Maharaj, 1997). Su importancia económica radica en la calidad de sus frutos, los cuales presentan propiedades alimenticias, medicinales, agroindustriales, y alto contenido de vitaminas A1, B1, B2 y C (Rodríguez, 2004). Por su importancia económica y alimentaria, se vienen realizando diferentes trabajos orientados a actualizar la tecnología de su cultivo en lo concerniente al manejo integrado y fertilización (OIRSA, 2002).
Entre las nuevas tecnologías empleadas en papaya, se encuentra el uso de microorganismos benéficos, que tienden a incrementar notablemente el crecimiento y desarrollo de este frutal (Sánchez, 2001; Hidalgo, 2012). Además, estos microorganismos pueden favorecer a las plantas permitiéndoles contrarrestar factores de estrés bióticos y abióticos (Hidalgo et al., 2014). Dentro de estos microorganismos, se han realizado experiencias independientes que evalúan el efecto de los hongos micorrícos arbusculares (HMA) y de hongos antagonistas como Trichoderma spp. (López, 2019; Quiñones, 2019). Es así que existen trabajos que han llegado a incrementar los diversos parámetros de crecimiento en diferentes etapas de desarrollo de C. papaya L. (Sánchez, 2001).
Los HMA forman una simbiosis biotrófica con las raíces de las plantas. En esta simbiosis se da un intercambio recíproco de nutrientes en el que el hongo recibe carbohidratos del hospedero, al tiempo que proporciona nutrientes minerales a través del micelio extrarradicular (Smith and Read, 2008). En C. papaya, los beneficios de los HMA incluyen aumento en el peso de la planta, aumento en la longitud de tallo y raíz, e índice de área foliar (Sharda & Rodriguez, 2009). Además, se ha observado que los árboles de papaya inoculados, reducen la necesidad de fertilización con P, hasta la etapa de producción (Mamatha et al., 2002). Además, estos microorganismos son capaces de disminuir el estrés y la pérdida de crecimiento sufrido por las plántulas luego de la aclimatación y a la vez promover el desarrollo vegetativo a nivel de invernadero (Hidalgo, 2012)
Por otra parte, la utilización de microorganismos antagonistas que se utilizan para disminuir el estrés producido por la presencia de organismos fitopatógenos a nivel de invernadero constituye una práctica creciente. Dentro de estos organismos, se encuentra Trichoderma viride. Este hongo es un saprófito capaz de estimular el crecimiento vegetal y rendimiento (Yadav et al., 2015). Esta acción podría estar ligada a la producción de metabolitos secundarios que tienen diversas propiedades sobre el desarrollo y defensa vegetal, tales como fitohormonas y antibióticos capaces de contrarrestar agentes patógenos (Contreras et al., 2016). En “papaya”, incluso se ha demostrado el efecto antagónico de Trichoderma spp., en contra de Fusarium oxysporum, responsable de la pudrición en plántulas (González et al., 2005).
Dado que las plantas de C. papaya tienden a ser más susceptibles al estrés por trasplante durante el primer mes, disminuyendo su crecimiento vegetativo y retardando su desarrollo, el presente trabajo tiene el objetivo de evaluar el efecto de la inoculación de los hongos benéficos Rhizophagus irregularis y Trichoderma viride en Carica papaya en condiciones de invernadero hasta los 28 días.
Materiales y métodos
Material biológico
Se emplearon plántulas clones de C. papaya obtenidas por micropropagación in vitro de meristemos en el Laboratorio de Biotecnología del Proyecto Especial Chavimochic. Estas fueron aclimatadas y enraizadas en invernadero durante 15 días, usando turba Kekkila comercial como sustrato. Las plántulas empleadas midieron entre 1,9 y 2,6 cm.
El inóculo de R. irregularis fue obtenido del producto comercial AEGIS Endo Gránulo, donado por la Empresa Agrotecnologías Naturales ATENS.
El inóculo de T. viride fue facilitado por el Laboratorio de Hongos Benéficos del Campamento San José del Proyecto Especial Chavimochic.
Tratamientos
Las cantidades de inóculo de ambos microorganismos fueron establecidas teniendo en cuenta las recomendaciones del personal técnico del P. E. Chavimochic para frutales en invernadero: 500 esporas de R. irregularis y 5x107 esporas de T. viride por planta. Los tratamientos estuvieron dados por las combinaciones de estas cantidades de inóculo, estableciéndose un control no inoculado, tal y como se indica en la Tabla 01. Los inóculos de ambos microorganismos fueron aplicados el mismo día al momento del trasplante a bolsas de plástico negro de 2 litros conteniendo arena y turba comercial esterilizada en una proporción 1:1. Los tratamientos fueron asignados de forma aleatoria a cada unidad experimental y las unidades experimentales se dispusieron en diseño completamente aleatorizado en el invernadero por 28 días.

Para realizar la inoculación se colocaron 5 g de inóculo sólido del hongo R. irregularis (100 propágulos.g-1 de inóculo) en contacto directo con las raíces de la plántula, mientras que el inóculo de T. viride fue agregado al sustrato en 50 ml una solución de 100 g inóculo por litro (concentración del inóculo: 106 esporas). Los 4 tratamientos fueron establecidos en un Diseño Completamente Aleatorio (DCA), con 5 repeticiones cada uno, cada unidad experimental constaba de una planta por bolsa. Las plantas fueron regadas con solución NPK 15-15-15 hasta los 28 días, momento en que se realizó la evaluación final.
Recolección y análisis de datos
Se evaluó longitud y diámetro de tallo, y número de hojas cada 7 días. El área foliar de la segunda hoja fue calculada a los 28 días, tomando como referencia la proporción gravimétrica (el área foliar de las siluetas se estimó tomando como referencia el peso de 18 cm2 de papel utilizado).
Los datos obtenidos fueron trabajados utilizando el software Minitab®, versión 19.1 (64 bit). Se realizó el análisis de varianza (ANVA) y el test de comparación múltiple de medias Tukey (α=0.05) para cada una de las variables consideradas. Los datos fueron organizados en gráficos resumen indicando la significancia con letras diferentes.
Resultados
La inoculación con R. irregularis y T. viride, originó cambios significativos en el crecimiento en invernadero tras 29 días de tratamiento (Fig. 2).

Fig. 2 C. papaya cultivada en condiciones de invernadero, a los 0 y 28 días de tratamiento con R. irregularis y T. viride.
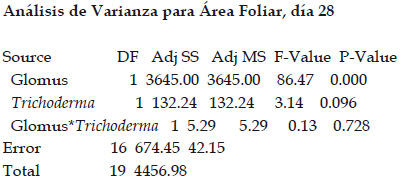
La longitud del tallo se incrementó de forma significativa desde el día 7 empleando el tratamiento con R. irregularis. Este incremento se mantuvo sobre los demás tratamientos a los 14, 21 y 28 días de tratamiento. Sin embargo, a los 28 días de tratamiento, se encontraron diferencias significativas entre el tratamiento con R. irregularis (4,32 cm) y el tratamiento con la coinoculación de R. irregularis y T. viride (3,54 cm), siendo ambos estadísticamente superiores a los tratamientos Control (2,70 cm) y con T. viride (2,62 cm) que a su vez fueron similares entre sí (Fig. 3).
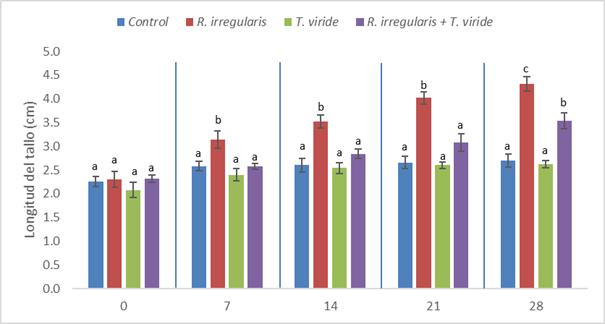
Fig. 3 Longitud de tallo de plántulas de C. papaya a los 0, 7, 14, 21 y 28 días de tratamiento. Letras diferentes indican diferencias significativas (p<0.05), según la prueba de comparación de medias múltiple de Tukey HSD. La línea vertical en la parte superior de cada barra representa el error estándar.
Al evaluarse el número de hojas (Fig. 4), no se observaron diferencias significativas entre los tratamientos hasta los 7 días. Posteriormente, al día 14 de tratamiento, se observa un incremento en el número de hojas en el tratamiento con R. irregularis (5,4 hojas.planta) con respecto al tratamiento control (3,4 hojas.planta). En tanto, en la misma fecha, se observaron valores intermedios en los tratamientos con T. viride (4 hojas.planta) y la coinoculación entre R. irregularis y T. viride (4,2 hojas.planta).
Para los días 21 y 28, el tratamiento con T. viride quedó rezagado junto al grupo control, mostrando los menores números de hojas (4,6 y 4,2 hojas.plantas al día 21 y 5 y 4,8 hojas.planta, respectivamente). Estos tratamientos, no mostraron diferencias significativas con respecto a la coinoculación de T. viride y R. irregularis (5,2 a los 21 días y 6 a los 28 días). Sin embargo, fueron estadísticamente inferiores al tratamiento con R. irregularis (6,6 hojas.planta al día 21 y 7,4 hojas.planta al día 28).
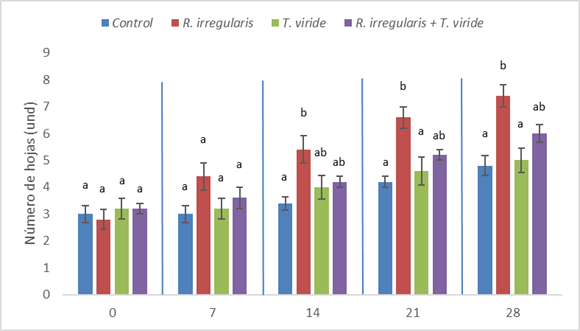
Fig. 4 Número de hojas de plántulas de C. papaya a los 0, 7, 14, 21 y 28 días de tratamiento. Letras diferentes indican diferencias significativas (p<0.05), según la prueba de comparación de medias múltiple de Tukey HSD. La línea vertical en la parte superior de cada barra representa el error estándar.
Los tratamientos indujeron diferencias significativas en el incremento en el diámetro de tallo a partir del día 14 de tratamiento, tal como se puede apreciar en la Fig. 5. En esta fecha, los tratamientos con R. irregularis y la coinoculación con R. irregularis y T. viride mostraron un diámetro de tallo (3,05 y 2,99 mm, respectivamente) mayor a las plantas del grupo control (2,38 mm). Hacia el día 21, el mayor diámetro de tallo se mantuvo en las plantas tratadas con R. irregularis (3,63 mm), las cuales mostraron un diámetro mayor que el de las plantas del grupo control (2.60 mm), en contraste con las plantas tratadas con T. viride y las coinoculadas con T. viride y R. irregularis (3,06 y 3,33 mm, respectivamente), que no mostraron diferencias con estos dos tratamientos.
Al llegar el día 28, las plántulas tratadas con R. irregularis (4,67 mm) mostraron un diámetro de tallo mayor que las plantas del grupo control (3,18 mm) y las inoculadas con T. viride (3,27 mm). Sin embargo, estas plantas mostraron valores de diámetro de tallo estadísticamente similares a las coinoculadas con T. viride y R. irregularis (4,05 mm).
Fig. 5 Diámetro de tallo de plántulas de C. papaya a los 0, 7, 14, 21 y 28 días de tratamiento. Letras diferentes indican diferencias significativas (p<0.05), según la prueba de comparación de medias múltiple de Tukey HSD. La línea vertical en la parte superior de cada barra representa el error estándar.
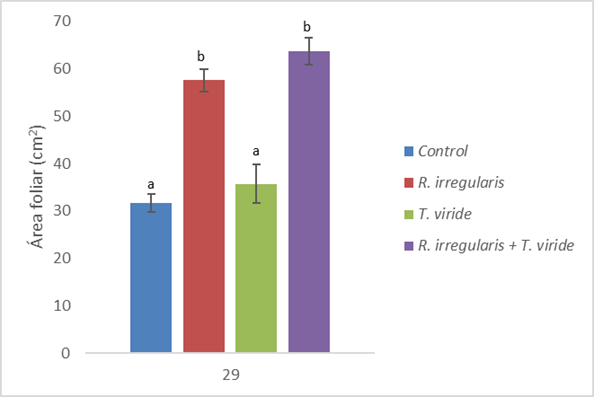
En cuanto al área foliar, se pueden apreciar las variaciones en el área foliar de C. papaya inducidas por los tratamientos a los 28 días. En la Fig. 6 se aprecia la ausencia de diferencias significativas entre el grupo control (31,63cm2) y el grupo de plantas inoculadas con T. viride (35,74cm2). En tanto, se aprecia un incremento inducido por la inoculación con R. irregularis (57,60cm2) con respecto al grupo control. El grupo tratado con las HMA mostró valores estadísticamente similares a las plántulas coinoculadas con T. viride y R. irregularis (63,77cm2).
Discusión
El uso adecuado de organismos benéficos, permite establecer sistemas autosostenibles, con reducidas aplicaciones de productos químicos y resultados económicamente rentables (Baar, 2008), lo cual es esencial en frutales, permitiendo reducir el tiempo de estancia de las plantas a nivel de invernadero. En particular, la utilización de R. irregularis y el hongo antagonista T. viride, son capaces de inducir efectos benéficos sobre el contenido de nutrientes de las plantas, su crecimiento vegetativo y su producción (Yadav et al., 2015). Esto coincide con los resultados obtenidos en el presente estudio, en el cual el uso de los tratamientos con R. irregularis y T. viride incrementaron las variables de crecimiento durante los primeros 28 días, ocasionando incrementos en la longitud y diámetro de tallo, número de hojas y área foliar de C. papaya. Tal como se puede apreciar en las figuras 2, 3, 4, 5 y 6, las variables de crecimiento evaluadas, se incrementaron principalmente en respuesta a la inoculación con R. irregularis. Este hongo es capaz de formar asociaciones biotróficas con las raíces de las plantas. Estas relaciones se caracterizan por un intercambio recíproco de nutrientes que permiten la supervivencia del hongo y mejoran la nutrición del hospedero, así como su tolerancia a diversos factores de estrés (Cornejo et al., 2017, Fuentes et al., 2016).
Es notorio el hecho de que el uso de T. viride por sí solo no tuvo un efecto positivo sobre las variables evaluadas, manteniendo valores de longitud y diámetro de tallo, número de hojas y área foliar similares al grupo control. Esta especie es capaz de establecer interacción con especies como Arabidopsis, “tomate”, “zapallo”, “arverja”, “canola” e incluso plantas leñosas (Vinale et al., 2008; Contreras et al., 2009; Samolski et al., 2012; Carrero et al., 2018). Además, pertenece a un grupo de hongos saprófitos, que son indispensables en la descomposición de la materia orgánica debido a la amplitud de su maquinaria enzimática (Mineki et al., 2015). Más específicamente, especies del género Trichoderma son conocidos como organismos promotores del crecimiento e inductores de la salud vegetal, principalmente por su producción de metabolitos secundarios activos biológicos, que incluyen fitohormonas y antibióticos contra fitopatógenos (Contreras et al., 2016). Este hongo antagonista establece relaciones antagónicas como micoparasitismo, antibiosis o competición en presencia de fitopatógenos y nemátodos, lo cual puede favorecer el desarrollo de las plantas (Lorito et al., 2010; Medeiros et al., 2017). Sin embargo, carece de mecanismos que permitan absorber los nutrientes presentes en el suelo e intercambiarlos con las plantas en una relación benéfica, en contraste con los HMA. Esto podría explicar la razón por la cual, la inoculación con T. viride no mostró efecto positivo con respecto al grupo control.
Al comparar los valores inducidos por R. irregularis por sí solo, se observa una ventaja con mayores valores de longitud y diámetro de tallo, así como mayor número de hojas en comparación a los obtenidos por la coinoculación de plantas con R. irregularis y T. viride. Eso podría ser explicado por la naturaleza de la interacción entre ambos microorganismos. Tal como lo reportaron Lagos et al. (2018), se puede observar inhibición mutua fuerte entre R. irregularis y T. viride, lo cual puede mitigar sus efectos promotores del crecimiento vegetal. Los mecanismos involucrados en la inhibición del crecimiento de R. irregularis por T. viride pueden estar ligados a la competición por nutrientes (Green et al., 1999), micoparasitismo o producción de compuestos secundarios antagónicos o cambios en la comunidad microbiana del suelo (Lagos et al., 2018).
En este sentido, se deben tener en cuenta los resultados obtenidos por Contreras et al. (2016) y Dehariya et al. (2015), quienes encontraron inhibición y promoción de la colonización de raíces por hongos micorrícicos en presencia de diferentes especies de Trichoderma. Esto denota que el resultado de las interacciones entre R. irregularis y Trichoderma es bastante complejo, dependiendo de las especies involucradas, así como de las condiciones ambientales. Lo cual hace necesario tomar en consideración la compatibilidad entre los organismos involucrados y la complejidad de sus interacciones en el desarrollo de complejos fúngicos con diferentes propósitos, como biofertilización, biocontrol o biorremediación (Lagos et al., 2018).
Por otra parte, la variación inducida por los tratamientos en cuanto al área foliar observados en las plántulas de C. papaya, muestra resultados discordantes con respecto a las otras variables tomadas en consideración. Tal como se observa en la Fig. 6, la coinoculación con R. irregularis y T. viride indujo una mayor área foliar en las plántulas de 28 días, en contraste con los resultados obtenidos en longitud y diámetro de tallo, así como en número de hojas. Esta variación puede deberse a la alteración de los niveles hormonales inducidos por el complejo micorrizas-Trichoderma, los cuales al interactuar ocasionan una atenuación de los niveles de ácido salicílico, ácido jazmónico y etileno, los cuales son capaces de alterar la localización de biomasa (Martínez et al., 2011). Esto podría potenciar los efectos benéficos sobre el crecimiento de hormonas como auxinas y citoquininas (Alcántara et al., 2019). El hecho de que este efecto no se pueda observar en presencia únicamente de T. viride, indica que la promoción del desarrollo de la lámina foliar requiere el establecimiento de una interacción entre los microorganismos evaluados en este estudio.
Conclusiones
El uso de organismos benéficos como R. irregularis y T. viride permite incrementar los parámetros de crecimiento de C. papaya (Caricaceae) en los primeros días de cultivo, desde los 7 hasta los 28 días en condiciones de invernadero.
La interacción de plántulas con R. irregularis indujo valores mayores de longitud y diámetro de tallo, así como número de hojas en C. papaya a los 7, 14, 21 y 28 días de tratamiento en condiciones de invernadero.
La utilización de T. viride no indujo cambios significativos en el crecimiento de plántulas de C. papaya a los 7, 14, 21 y 28 días de tratamiento en condiciones de invernadero.
La coinoculación de plántulas de C. papaya con T. viride y R. irregularis permitió incrementar los parámetros de crecimiento evaluados, mostrando mayor efecto en el incremento del área foliar de las plántulas a los 7, 14, 21 y 28 días de tratamiento en condiciones de invernadero.