











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
El aumento de sales en los suelos es una de las más serias amenazas medioambientales para la supervivencia de las plantas (Munns, 2005; Ashraf, et al., 2003). En muchas áreas del mundo dedicadas a la agricultura, la obtención de buenos rendimientos, así como también la capacidad de cultivar una amplia variedad de especies, tiene muchas restricciones debido a la salinización de los suelos (Tanwar, 2003; Goykovic, 2007). Este problema junto con la sequía, constituyen los factores abióticos que producen mayor descenso en el rendimiento de los cultivos (Reigosa et al., 2004).
La mayoría de los daños provocados por salinidad son debidos por las sales de sodio (Pessarakli, 2002; Jenks et al., 2007), principalmente, NaCl (Miranda, 2011), sal que se encuentra frecuentemente en terrenos de cultivo, por ello, muchos estudios han sido orientados al entendimiento de los mecanismos involucrados en la respuesta de las plantas al estrés producido por NaCl (Casierra et al., 2013; Taiz et al., 2007; Yokoi et al., 2002).
Los efectos del NaCl sobre la fisiología de las plantas han sido clasificados como de toxicidad, osmóticos y nutricionales (Jenks et al., 2007). El primero constituye un efecto primario debido a las sales, mientras que los siguientes son efectos secundarios inducidos por el estrés salino (Coca et al., 2012; Reigosa et al., 2004). Ante estas situaciones, las plantas desarrollan mecanismos de respuestas morfológicas, fisiológicas y bioquímicas. Estas respuestas varían ampliamente dependiendo del genotipo y del estadio de desarrollo de la planta. Mientras algunas especies presentan elevada tolerancia a la salinidad otras son altamente susceptibles (Taiz et al., 2007; Zhu, 2003).
El “tomate” (Solanum lycopersicum L.), es una hortaliza ampliamente distribuida alrededor del mundo, ocupando el segundo lugar en importancia mundial teniendo en cuenta su valor económico y nutricional (Giaconi, 1995; Casanova, 2007). Posee un alto contenido de B- carotenos (licopeno), vitamina A, vitamina C (ácido ascórbico), minerales (calcio, hierro, fósforo y potasio) y aminoácidos (tiamina y niacina) (Willcox et al., 2003).). Además, se le han atribuido muchas propiedades medicinales tales como anticancerígeno, antiséptico, analgésico, sedante; además se reportan propiedades antidiabéticas (Brecht et al., 2004; Ramandeep, 2005; Juroszek et al.,2009).
El cultivo del “tomate” en áreas con problemas de salinidad provoca en las plantas un sinnúmero de efectos fisiológicos, morfológicos y bioquímicos, entre los cuales se encuentran cambios cuantitativos y cualitativos en la síntesis de proteínas por cambios en la expresión de genes, entre otros (Goykovic et al., 2007). Así, las plantas sometidas a estrés salino pueden sintetizar proteínas reguladoras, dentro de las cuales se encuentran los factores de transcripción, proteínas quinasas, fosfolipasas. Además, pueden producir proteínas de respuesta, como proteínas LEA (Late Embryogenesis Abundant o dehidrinas), enzimas para la biosíntesis de sustancias osmoprotectoras, proteínas de choque térmico (HSPs, heat shock proteins) (chaperonas), acuaporinas, proteasas, enzimas detoxificadoras (Reigosa et al., 2004; Taiz et al., 2007).
Para atenuar el efecto adverso de las sales en el rendimiento de los cultivos de interés agrícola como el tomate, existen una serie de prácticas agronómicas posibles de implementar como el uso de ciertos microorganismos, particularmente bacterias y hongos benéficos. Uno de los microorganismos más comunes es Rhizophagus irregularis, este hongo no solo maximiza la captación de nutrientes, sino que también le permite a su vez superar situaciones de estrés biótico y abiótico en las plantas (Tapia, 2003; Rouhrazi et al., 2016).
Los suelos de la costa peruana son afectados por NaCl, el cual restringe el crecimiento y productividad del tomate, planta que modifica significativamente su expresión génica en respuesta a una serie de cambios a nivel molecular. Por esta razón, el objetivo del presente trabajo fue evaluar el efecto del Rhizophagus irregularis y NaCl en el patrón electroforético de proteínas solubles totales de tomate (Solanum lycopersicum), cuantificar y separar mediante electroforesis SDS-PAGE las proteínas solubles totales en hojas y raíces de tomate por efecto de la inoculación con 0, 1.5 y 3 g de Rhizophagus irregularis y tratadas con 0, 100 y 200 mM de NaCl.
Materiales y métodos
Material biológico
Se utilizaron plántulas de tomate (Solanum lycopersicum L.) var. Rio Grande de 15 días de germinadas obtenidas a partir de semillas certificadas, las cuales fueron colocadas en un sistema de germinación que consistió en recipientes de tecnopor con papel toalla absorbente como base. Para la micorrización se utilizó el inóculo de Rhizophagus irregularis, el cual se obtuvo del producto comercial AEGIS Micro Gránulo, donado por la empresa ATENS Agrotecnologías Naturales, S.L. Tarragona, España.
Inoculación de plantas de tomate con Rhizophagus irregularis y aplicación de NaCl
Se seleccionaron 90 plántulas en base a su uniformidad en tamaño, las cuales fueron aleatoriamente distribuidas en 9 grupos. El sustrato estuvo constituido por arena de río y musgo. La arena de río fue cernida con un tamiz para homogeneizar su textura, después fue lavada con agua corriente por 8 horas para eliminar impurezas, tras lo cual se esterilizó en la estufa a 200° C, durante 3 horas, para eliminar los agentes patógenos. Luego, la arena fue mezclada con el musgo en proporción 1:1. El sustrato obtenido fue colocado en recipientes de polietileno de 1000 ml de capacidad.
Para realizar la inoculación, en los recipientes con sustrato se hizo un orificio de aproximadamente 3 cm. de diámetro, para recibir a las plántulas. A continuación, se agregó a dicho orificio el inóculo de Rhizophagus irregularis, de manera que las raíces estuvieron en contacto con el HMA.
Bajo condiciones de invernadero a una temperatura promedio de 25 ± 2°C y humedad relativa de 75 ± 2%, las plantas crecieron por 15 días y fueron regadas con la solución nutritiva La Molina de 2 a 3 veces por semana. Transcurrido ese tiempo, las plantas fueron sometidas a estrés salino, para lo cual éstas fueron regadas con solución nutritiva y NaCl que generen concentraciones de 0, 100 y 200 mM por un periodo de 30 días (Figura 1).

Obtención y cuantificación de proteínas solubles totales
Se pesó 5 g de tejido fresco de hojas y raíces. Luego, por separado, los tejidos se trituraron con nitrógeno líquido en un mortero de porcelana, utilizando como buffer de extracción fosfato de sodio 50 mM y pH 7. El homogenizado obtenido se centrifugó a 10 000 g durante 10 minutos a 4°C; enseguida el sobrenadante se utilizó para cuantificar proteínas. Se siguió el método de Bradford (1976), para lo cual, se tomó 10 ul del extracto y se llevó el volumen final a 1 ml con el reactivo de Bradford, luego se medió la absorbancia en un espectrofotómetro Genesys 6 a una longitud de onda de 595 nm. Para determinar la cantidad de proteínas se utilizó BSA como estándar en la curva de calibración.
Electroforesis de las proteínas
Para realizar la electroforesis de las proteínas se pesó 5 g de tejido fresco de raíces y de hojas, y se siguieron todos los pasos mencionados anteriormente para la obtención de proteínas. Las proteínas fueron separadas mediante electroforesis en geles de poliacrilamida SDS - PAGE (dodecilsulfato sódico-polyacrylamide gel electrophoresis), de acuerdo a Laemmli (1970). Se utilizó geles de Acrilamida/Bis al 30%.
Los geles fueron cargados con 20 ul del extracto y el corrido electroforético se realizó utilizando una cámara Mini-Protean III de Bio-Rad, siguiendo las especificaciones recomendadas por el Manual (Figura 2).

La tinción de los geles se efectuó sumergiéndolos en una solución de azul de Coomassie, manteniéndolos en agitación constante por 40 a 50 minutos. Luego de la decoloración, los geles fueron fotografiados para el análisis respectivo.
Análisis de datos
Una vez obtenidos los resultados, fueron ordenados en tablas y gráficos y se analizaron estadísticamente, para determinar si existen diferencias significativas entre tratamientos, mediante el análisis de varianza y la prueba de comparación múltiple Tukey HSD.
Las bandas reveladas en los geles fueron analizadas para encontrar su peso molecular mediante interpolación de los pesos moleculares de proteínas conocidas, utilizando como patrón una mezcla de proteínas de peso molecular conocido.
Resultados
El contenido de proteínas solubles totales tanto en hojas, como en raíces de tomate (Solanum lycopersicum) tratadas con 0, 100 y 200 mM de NaCl se incrementó de manera general, conforme aumentó la cantidad de inóculo de Rhizophagus irregularis, al cabo de 30 días.
En la figura 3, se presenta el contenido de proteínas solubles totales en hojas, según tratamiento, se observa que en los tratamientos con 0mM de NaCl, existe un aumento significativo (p<0,05) en el contenido de proteínas solubles totales cuando se agregó el hongo, observándose valores de 0.650 y 0.774 mg/g p.f. respectivamente, en los tratamientos con 1.5 y 3 g de inóculo.
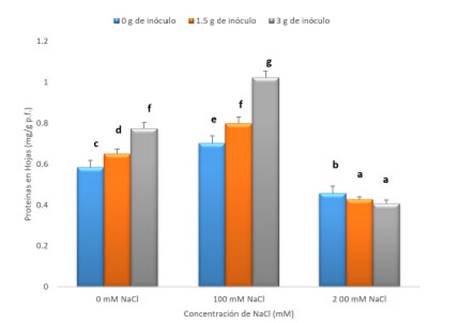
Fig. 3 Contenido de proteínas solubles totales (mg/g p.f) en hojas de tomate (Solanum lycopersicum L). tratadas con Rhizophagus irregularis Schenk & Smith (1, 1.5 y 3 g de inóculo) y expuestas a concentraciones de NaCl (0, 100 y 200 mM) por 30 días de tratamiento. Las barras verticales indican el error estándar. Las letras diferentes indican diferencias debidas al tratamiento según Tukey HDS (n=3).
La misma tendencia se observa en las plantas expuestas a 100 mM de NaCl, en las que se encontró que los tratamientos con 1.5 y 3 g de inóculo, indujeron los valores más altos (0.797 y 1.023 mg/g p.f. respectivamente), siendo estas plantas las que tuvieron el más elevado contenido de proteínas solubles totales.
En contraste, las plantas expuestas a 200 mM de NaCl y tratadas con el inóculo, mostraron los menores valores de proteínas solubles totales (0.427 y 0.407 mg/g p.f. respectivamente) al ser comparados con el control, independientemente de la adición del hongo.
La Figura 4, se presenta el patrón electroforético en hojas de tomate (Solanum lycopersicum L.) tratadas con Glomus intraradices (1, 1.5 y 3 g de inóculo) y expuestas a concentraciones de NaCl (0, 100 y 200 mM), por 30 días. Las plantas tratadas con 0 mM de NaCl y cantidades de inóculo mostraron cambios de intensidad en algunas bandas proteicas; así, las bandas de 154, 92, 53 y 30 kDa disminuyen de intensidad cuando se agrega 1.5 g de inóculo, mientras que las bandas de 154 y 92 kDa con 3 g de inóculo aumentan.

Fig. 4 Patrón electroforético de proteínas solubles totales en hojas de tomate (Solanum lycopersicum L). tratadas con Rhizophagus irregularis Schenk & Smith (1, 1.5 y 3 g de inóculo) y expuestas a concentraciones de NaCl (0, 100 y 200 mM) por 30 días de tratamiento. A la izquierda se observan los pesos moleculares de las proteínas en kDa. A la derecha((= , aparición de una proteína, (= desaparición de proteína.
En las plantas tratadas con 100 mM de NaCl, se observa una disminución de las intensidades de las bandas de 154, 88, 50 y 30 kDa cuando se agrega 3 g de inóculo. En las plantas tratadas con 200 mM de NaCl, se observa un aumento en la intensidad de las bandas proteicas independientemente de la cantidad de inóculo. Asimismo, se observa la aparición de proteínas de 111 kDa cuando se agrega 1.5 y 3 g de inóculo. También, se observa la desaparición de una proteína de 50 kDa cuando se agrega 3 g de inóculo.
En la Figura 5, se presenta el contenido de proteínas solubles totales (mg/g p.f) en raíces de tomate (Solanum lycopersicum L.), según tratamiento. Esta gráfica muestra que el contenido de proteínas en este órgano disminuye conforme incrementa la concentración de NaCl, tanto para plantas inoculadas, como aquellas que no fueron tratadas con el organismo fúngico.
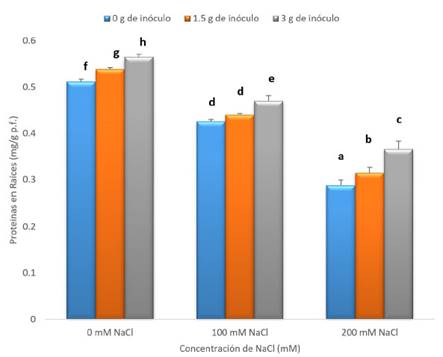
Fig. 5 Contenido de proteínas solubles totales (mg/g p.f) en raíces de tomate (Solanum lycopersicum L.) tratadas con Rhizophagus irregularis Schenk & Smith (1, 1.5 y 3 g de inóculo) y expuestas a concentraciones de NaCl (0, 100 y 200 mM) por 30 días de tratamiento. Las barras verticales indican el error estándar. Las letras diferentes indican diferencias debidas al tratamiento según Tukey HDS (n=3).
Las plantas tratadas con 0mM de NaCl, muestran los valores más elevados de proteínas solubles totales con respecto a los demás tratamientos. En este grupo, se observa un aumento significativo (p<0,05) en los resultados cuando se agregó 1.5 y 3 g de inóculo, encontrándose valores de 0.538 y 0.565 mg/g p.f. respectivamente.
En las plantas expuestas a 100 mM de NaCl, se encontró que los tratamientos con 1.5 y 3 g de inóculo, indujeron un aumento, con valores de 0.439 y 0.470 mg/g p.f. de proteínas solubles totales respectivamente. Sin embargo, estos valores son menores, al ser comparados con el grupo control.
La misma tendencia muestran los tratamientos con 200 mM de NaCl e inóculo, en las que mayores cantidades de inóculo indujeron un aumento en el contenido de proteínas solubles totales. Estas plantas, a su vez, mostraron los menores valores de proteínas solubles totales (0.288, 0.315 y 0.366 mg/g p.f. respectivamente) al ser comparados con el control.
En la Figura 6, se muestra el patrón electroforético de raíces de Solanum lycopersicum L. tratadas con Rhizophagus irregularis (1, 1.5 y 3 g de inóculo) y expuestas a concentraciones de NaCl (0, 100 y 200 mM), por 30 días; el cual reveló la aparición de una proteína de 73 kDa cuando son tratadas con 200 mM de NaCl, independiente de la cantidad de inóculo. Asimismo, la banda de peso molecular 73 kDa se presenta en las plantas tratadas con 100 NaCl y 3 g de inóculo. También, se observa que las bandas proteicas en general disminuyen de intensidad. Sin embargo, las bandas de pesos moleculares 96 kDa aumentan ligeramente de intensidad cuando son expuestos a 200 mM de NaCl. Las bandas con los pesos moleculares de 146 kDa no presentan cambios entre los tratamientos.

Fig. 6 Patrón electroforético de proteínas solubles totales en raíces de tomate (Solanum lycopersicum L.), tratadas con Rhizophagus irregularis Schenk & Smith (1, 1.5 y 3 g de inóculo) y expuestas a concentraciones de NaCl (0, 100 y 200 mM) por 30 días de tratamiento. A la izquierda((= se observan los pesos moleculares de las proteínas en kDa. A la derecha, aparición de una proteína.
Discusión
Las plantas que crecen en suelos con NaCl, suelen presentar desbalance osmótico, desequilibrio nutricional y, principalmente toxicidad iónica, estos efectos involucran respuestas fisiológicas y bioquímicas (Bartels et al., 2005). Entre las principales respuestas de las plantas al estrés salino está la modificación de su expresión génica, que puede inducir la acumulación o agotamiento de ciertos metabolitos, alteraciones en el comportamiento de muchas enzimas, cambios globales en la síntesis de proteínas y en especial la síntesis de nuevos conjuntos de proteínas (Lei et al., 2009; Lesmes et al., 2007; Taiz et al., 2007).
En el presente trabajo, se observó que, en hojas de tomate (Solanum lycopersicum) tratadas con 0 y 100 mM de NaCl, el contenido de proteínas solubles totales aumenta, conforme se incrementan las cantidades de inóculo de R. irregularis (Fig. 3). Las proteínas sintetizadas bajo estrés salino cumplen un papel importante en la tolerancia o adaptación de las plantas a este factor de estrés (Casierra et al., 2013). Sin embargo, el mecanismo general de cómo estas proteínas podrían proporcionar la adaptación no es entendido en su totalidad. Bajo estrés salino, junto con los aminoácidos y compuestos nitrogenados solubles, actúan como componentes de un mecanismo de tolerancia a la sal (Fita et al., 2015). Estos pueden funcionar también como solutos compatibles citoplasmáticos en el ajuste osmótico, con el fin de igualar el potencial osmótico del citoplasma y las vacuolas en condiciones adversas de salinidad (Casierra et al., 2013; Karakas et al., 2016). Por otro lado, un incremento inicial en las proteínas solubles totales durante el estrés podría deberse a la expresión de nuevas proteínas; pero su disminución cuando se incrementa la intensidad de los factores de estrés, se debería a una severa reducción de los materiales necesarios para la síntesis proteica (Mohammadkhani, 2008; Fita et al., 2015).
En plantas sometidas al estrés salino, la síntesis de proteínas de respuesta a este factor es inducida por la activación de rutas dependientes e independientes del ABA, donde la transcripción es controlada por la interacción de proteínas reguladoras (factores de transcripción) y secuencias reguladoras específicas de los promotores de los genes (ABRE y DREA) (Chinnusamy et al., 2005; Taiz et al., 2007). Así, en plantas de Arabidopsis thaliana el factor de transcripción DREB1A, permite activar la expresión de genes involucrados en la tolerancia al estrés, mejorando la tolerancia a la sequía, heladas y salinidad (Kasuga et al., 1999; Fita et al., 2015). También en plantas transgénicas de Solanum lycopersicum, se ha descrito el factor de transcripción, HDZIPs (Homeodominio-Leucina Zipper Proteins), que participa en la activación de genes que mejoran la tolerancia al estrés salino (Bartels et al., 2005).
Asimismo, diversas investigaciones señalan que los hongos formadores de micorrizas también tienen la habilidad de proteger a las plantas del estrés provocado por la salinidad, pero los mecanismos que ocurren no están muy claros. De esta forma, el estrés por NaCl, conduce a la expresión diferencial de la información genética, produciendo cambios en los productos génicos, incluyendo mRNA y proteínas (Coca et al., 2012; Casierra et al., 2013). Es así como aumenta la síntesis de nuevas proteínas en plantas estresadas y disminuye el nivel de centenares de proteínas existentes. Estas proteínas específicas de estrés, posiblemente dotan a las plantas con la capacidad de adaptarse al estrés medioambiental por medio de ajustes fisiológicos y bioquímicos (Lesmes et al., 2007).
Según Boucher et al. (1999), altas concentraciones de proteínas en maíz han sido asociadas con un rol adaptativo de las micorrizas arbusculares en la reducción de la degradación proteica bajo condiciones de estrés y a la producción de proteínas llamadas micorrizinas, las cuales pueden ser inducidas por la asociación con Glomus spp., en respuesta a la infección con el hongo, pero su función específica aún se desconoce. No obstante, los pocos datos disponibles indican que estos hongos contribuyen con el aumento del contenido de proteínas de estrés, las cuales forman parte de la respuesta al incremento moderado de NaCl (Aboul, 2011; Cristobal et al., 2010). En hojas de Ocimum basilicum tratadas con G. mosseae y G. intraradices expuestas a 0, 75 y 150 de NaCl, el contenido de proteínas solubles totales aumenta (Enteshari et al., 2012; Rouhrazi et al., 2016). En plantas de Zea mays tratadas con 0, 25, 50 y 100 de NaCl e inoculadas con Glomus spp. se encontró un aumento de proteínas solubles totales en el tejido foliar (Boucher et al., 1999; Bhattarai et al., 2011).
En este trabajo se observó que en raíces de S. lycopersicum “tomate”, el contenido de proteínas solubles totales disminuye tanto en plantas no inoculadas como en las inoculadas cuando se incrementa la concentración (Fig. 5). Este descenso observado puede ser el resultado de la inactivación de algunas enzimas o la inhibición de la síntesis proteica por el efecto primario del NaCl, toxicidad y/o el desequilibrio osmótico, principalmente en este nivel (Hassanein, 2004; Gomathi, 2006). Al respecto algunos investigadores refieren que la salinidad presenta un efecto especifico dependiente de la toxicidad de los iones en solución; y específicamente la toxicidad por NaCl no sólo se debe a efectos tóxicos del Na+ y del Cl- en el citosol, sino también por que la homeostasis del K+ es alterada, ocasionando la reducción de la relación K+/ Na+ en el citoplasma, posiblemente por la habilidad del Na+ de competir por los sitios de unión del K+(Bartels, 2005; Taiz, 2007; Zhu, 2002; Casierra et al., 2013). Generalmente, el Na empieza la inhibición de muchas enzimas a concentraciones próximas a 100 mM, la concentración a la cual el Cl- se vuelve tóxico aún no está bien definido, pero es probable que este en el mismo rango que el Na+ (Munns, 2005; Salazar, 2007). Situaciones similares podrían explicar esta disminución del contenido de proteínas solubles totales por efecto del NaCl, encontrado en este trabajo.
En el análisis del patrón electroforético de hojas de S. lycopersicum (Fig. 4), se encontró que las bandas que representan a una proteína de peso molecular 50 kDa desaparece y que las intensidades de algunas bandas proteicas disminuyeron (154, 92, 53 y 30 kDa), por efecto del NaCl. Estos cambios en las proteínas pueden estar relacionados con la respuesta y/o adaptación de S. lycopersicum al NaCl. Este comportamiento ha sido observado en otras especies, en Hordeum vulgare la intensidad de las bandas proteicas de peso molecular 176, 160, 147, 134 y 130 kDa aumentó y de las bandas de 55 y 15 kDa disminuyó por efecto del NaCl (50 y 100 mM) (Todorova et al., 1992). En Triticum durum expuestas a NaCl 200 mM, muestran que cuatro proteínas (24.8 y 27.9 kDa) se pierden por completo (Yildiz, 2006). En Bruguiera parviflora tratadas con diferentes concentraciones de NaCl (100, 200 y 400 mM), se encontró que la intensidad de varias bandas de peso molecular 17, 23, 32, y 34 kDa disminuyen como resultado del tratamiento con NaCl (Parida et al., 2004).
Al analizar el patrón electroforético de raíces (Fig. 6), se encontró que la intensidad de las bandas proteicas de peso molecular 146, 96 y 73 kDa disminuyeron debido al NaCl, respuestas similares se encontró en raíces de Zea mays, que al ser tratadas con NaCl, se indujo la acumulación de algunos polipéptidos con peso molecular de 148, 122.5, 118, 61, 51, 39 y 29 kDa (Tamas et al., 2001; Zorb et al., 2004). Estos cambios, estarían involucrados en la respuesta de S. lycopersicum frente al estrés salino, a nivel de raíz.
Conclusiones
La inoculación con Rhizophagus irregularis Schenck & Smith y el NaCl en hojas de “tomate” (Solanum lycopersicum), causaron un aumento significativo en el contenido de proteínas solubles totales, siendo el tratamiento de 100 mM de NaCl y 3 g de inóculo las que tuvieron el más elevado contenido de proteínas solubles totales.
En raíces, el contenido de proteínas solubles totales disminuye conforme incrementa la concentración de NaCl, tanto para plantas inoculadas, como aquellas que no fueron tratadas con el organismo fúngico.
Rhizophagus irregularis y el NaCl indujeron cambios en el patrón electroforético de proteínas solubles totales en hojas y raíces de tomate (Solanum lycopersicum L.) a 200 mM de NaCl con 1,5 y 3 g de inóculo.

