











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
En la naturaleza, los metales pesados pueden estar presentes como componentes del suelo, a manera de sales minerales y de otros compuestos. Su degradación es difícil de realizarse en forma natural o biológica por las diversas funciones metabólicas dentro de los mismos organismos (Abollino et al., 2002).
Los metales pesados como el cobre (Cu), zinc (Zn) y selenio (Se) son de gran importancia porque mantienen un adecuado metabolismo en los organismos vivos. No obstante, en altas concentraciones tienen la posibilidad de generar contaminación del agua potable, de la atmósfera alrededor de las fuentes generadoras de emisiones y en romper las cadenas tróficas (Kabata-Pendias, 2000). Entre los metales pesados más importantes está el cobre, debido a que es un metal primordial en el crecimiento y desarrollo regular de las plantas. Su capacidad de retención o fitoabsorción en los tejidos vegetales es de 10 microgramos, tiene participación en varios procedimientos bioquímicos de la respiración, fotosíntesis como respuesta al estrés oxidativo; actúan en el metabolismo de síntesis de hormonas y formación de la pared celular, asimismo es cofactor de algunas enzimas como citocromo oxidasa, superóxido dismutasa, amino oxidasa, oxidasa, polifenol, entre otros; poseen un papel primordial en la señalización y transporte de proteínas (Yruela, 2005).
El cobre es incorporado a través de las plantas como iones divalentes en suelos con presencia de oxígeno y como iones monovalentes en suelos con bajos niveles de oxígeno. La absorción se inicia en la epidermis de la raíz y su desplazamiento continúa hacia la endodermis por la vía apoplástica, finalmente la translocación del cobre por medio de la raíz hacia las hojas, en el xilema, el cual se transporta de forma pasiva por medio de la transpiración (León & Sepúlveda, 2012). En consecuencia, la toxicidad del cobre ocurre de manera natural en algunos suelos que contengan niveles altos, causado por la actividad del hombre. Es necesario resaltar que los metales entre ellos, el cobre estimulan en la formación de radicales hidroxilos que causan estrés en las plantas y a consecuencia de ello incrementan la respuesta hacia los antioxidantes (Yruela, 2005).
Es evidente la presencia de ciertos mecanismos de defensa contra el cobre en las plantas, por ejemplo, la relación existente entre las raíces con hongos micorrícicos. Se realizó un experimento al introducir a micorrizas arbusculares en dos plántulas de Populus sp “álamo” para interacción simbiótica, luego se sembraron en suelos contaminados con presencia de cobre y en suelos ausente del metal cobre; se evaluó los efectos en su crecimiento, determinando qué función cumplen los hongos micorrícicos en el alivio del estrés oxidativo. Se concluyó que ambas plantas poseían ciertas diferencias respecto a la contaminación por cobre y a la simbiosis con micorrizas, lo cual confirma que se podría usar plantas que presentan interacción interespecífica simbiótica, para futuros estudios de fitorremediación en suelos contaminados (Castiglione et al., 2007).
La fitoextracción es dependiente de ciertos componentes, primordialmente de la disponibilidad del metal en el ambiente para su absorción a través de la raíz y de la función en su transporte de sustancias a través del xilema y la biocumulación de los metales en los brotes. Su disponibilidad estaría controlada por microorganismos que catalizan la reacción de reducción y oxidación (redox), movilizan los metales presentes en el suelo para ser absorbidos por las raíces. Las plantas liberan ciertos compuestos orgánicos que sirven como nutrientes para los microorganismos, perjudicando indirectamente y a la movilidad de los cationes Cu+2 (Ducic & Polle, 2005).
Amarowicz et al. (2009) evaluaron la concentración y acumulación de sulfato de cobre, además la oxidación lipídica y el contenido total de componentes fenólicos encontrados en raíces de Lens culinars Medic. Las plantas fueron expuestas a una concentración de 0,5 mM de cobre durante 3 y 5 días en total oscuridad. Con este estudio se demostró que en concentración alta de cobre produce una disminución en la producción de ambos cultivares de lenteja. Concluyendo así, que el cobre induce al daño oxidativo y que además en un grupo causó una disminución en la producción de compuestos fenólicos, sin embargo aumento la peroxidación lipídica, lo que indica que el estrés oxidativo se generó por el incremento en la concentración de cobre que ha sido mayor a la capacidad antioxidante.
Ante esta problemática se realizaron diferentes estudios, en las cuales existió un cierto interés por el desarrollo de diversas pruebas biológicas que permitan identificar los efectos o daños citotóxicos y genotóxicos generados por los contaminantes en diferentes ecosistemas (Leme & Marín-Morales, 2009). Según Fiskesjö (1985 y 1997), los organismos biológicos poseen la capacidad de manifestar cambios cuando están expuestos a lo largo de un cierto tiempo a algún contaminante por medio de respuestas singulares como (reacciones morfológicas, etológicas, fisiológicas y bioquímicas), o de grupo (cambios en la composición y perturbaciones en la cadena trófica, entre los principales), las especies bioindicadores como su nombre lo indica, se permitiría identificar la calidad de las aguas residuales y/o efluentes, indirectamente caracterizando cualitativamente la toxicidad de ciertos elementos químicos, los cuales se esperarían que se encuentren dentro de los límites máximos permisibles o por debajo de los mismos. Con el pasar del tiempo, los ensayos o test biológicos se están convirtiendo en herramientas muy necesarias para evaluar los efectos que causan las sustancias tóxicas en el ecosistema.
En las últimas décadas, se realizaron diferentes ensayos usando diversas plantas como organismos indicadores, lo cual ha funcionado perfectamente como importantes herramientas en el monitoreo ambiental. En Cuba, cuentan con varios proyectos ejecutados utilizando plantas en la evaluación de toxicidad (Jerez et al., 2000). Las plantas superiores por ser organismos eucarióticos son muy utilizadas como bioindicadores y en consecuencia la posibilidad de tener estudios comparativos con otras especies de flora, por su susceptibles a cualquier cambio ambiental que otros tipos de ensayos evaluados (Dutka, 1989), siendo las plantas bioindicadores un factor y una herramienta de gran precisión, a diferencia de los parámetros físicos y químicos, en especial en los trabajos de investigación de manejo de recursos y riesgo ambiental (Fiskesjö, 1993; Gopalan, 1999).
El uso en la industria de iones metálicos (Hg, Cu, Ni, Cd, Be, Al, Mn, Li), metales pesados (Cd2+y Zn2+), colorantes textiles, productos desinfectantes (NaClO, CIO2 y CH3-CO-COOH) pesticidas, gran cantidad de contaminantes; todos ellos se caracterizan por ser sustancias clastogénicas (Leme & Marín-Morales, 2009), sería de gran relevancia el dirigir diversas investigaciones sobre evaluación, determinación de los efectos de los diversos contaminantes en los cuerpos de agua, los cuales finalmente van a ser parte del uso del recurso para el consumo humano, tanto para aguas, aguas superficiales, aguas subterráneas y efluentes domésticos. En ese sentido, La Real Academia Sueca de las Ciencias y el GENE - TOX PROGRAM, programa Internacional de Bioensayos Vegetales, ejecutan investigaciones al respecto en el uso de bioindicadores fundamentalmente en los estudios de bioensayos y ensayos relacionados a citotoxicidad y genotoxicidad de diferentes sustancias (Firbas, 2011).
Las pruebas que más se han usado en un breve lapso de tiempo para la evaluación de toxicidad ambiental se encuentra, el bioensayo con semillas de Lactuca sativa L. “lechuga” y de Solanum lycopersicum L. “tomate”, donde se evaluaron los efectos citotóxicos usando un solo compuesto o sustancia tóxica y proporciones de mezcla de tóxicos durante el proceso de germinación y desarrollo de las plántulas en los primeros días de crecimiento (Wang, 1991). Estudios realizados en bioensayos permitieron hacer una evaluación sobre el riesgo toxigénico de una o varias sustancias. Levan (1938) demostró los efectos citogenéticos de colchicina en el ciclo celular de meristemos radiculares de Allium cepa, asimismo Larroudé et al. (2016) encontró que A. cepa “cebolla” es posible el uso como bioindicador en la evaluación de calidad del agua. Cuando el bulbo se rehidrata se produce un incremento en el crecimiento celular permitiendo que se genere una elongación en sus raíces. Sin embargo, cuando el procedimiento de hidratación del bulbo se realiza en presencia de sustancias tóxicas, afecta la división de la mitosis celular a nivel de los meristemos radiculares produciendo inhibición, retardo o destrucción de células meristemáticas en mitosis. Estas alteraciones por lo regular dificultan el crecimiento adecuado de la raíz. Los efectos pueden hallarse de forma indirecta, a través de una comparación en el crecimiento de las raíces de los bulbos que estuvieron en contacto con el compuesto tóxico y con los que no estuvieron en contacto. El Test Allium es un ensayo modelo realizado en una especie, donde se evalúa el efecto producido por una sustancia o solución de interés en el desarrollo y crecimiento de las raicillas y en el material genético ADN (Díaz et al., 2004).
Según Leme & Marín (2009) las aberraciones cromosómicas son daños producidos a nivel celular, que se caracterizan por presentar modificaciones en la distribución y en la cantidad total de los cromosomas. Las aberraciones o alteraciones cromosómicas son probablemente inducidas por diversos factores, tales como la ruptura del ADN, bloqueo o afectación de la síntesis de ADN y la replicación del material genético, observando daño a este nivel del ADN. Las alteraciones presentes con más frecuencia son los puentes anafásicos múltiples y únicos, cuyos cromosomas muestran un estiramiento que en ciertos puntos los hace parecer tan delgados y pegajosos.
Se determinaron altos niveles de toxicidad en cobre en diferentes clases de suelos, como consecuencia de la actividad antropogénica. El cobre como metal pesado de transición cataliza la formación de radicales hidroxilos, y como resultado causan estrés oxidativo en las plantas (Yruela, 2005). Por otro lado, la relevancia del Test Allium radica por ser un ensayo modelo de un organismo vivo, mediante el cual se puede estimar los daños producidos por una sustancia tóxica, elemento o solución de interés en el desarrollo y crecimiento de las raíces y en el ADN (Díaz et al., 2004). Se ha sistematizado el Test Allium como un protocolo válido para determinar anomalías por la Organización Mundial de la Salud (OMS), el uso como bioindicador de toxicidad de ambientes contaminados, es muy significativa debido a que determina el daño citotóxico que se produce a nivel celular en los seres vivos. Es ese sentido, los resultados del presente trabajo, servirán en un futuro para la elaboración de medidas frente a la problemática que afecta al campo agrícola, A. cepa es una planta de gran importancia y de un gran valor nutricional, el cual podría ser alterado por contaminantes y afectar la salud humana.
Esta investigación tuvo por objetivos evaluar la citotoxicidad de cobre en Allium cepa L. “cebolla” a concentraciones de 500 µg/l, 600 µg/l, 700 µg/l y 800 µg/l y establecer el índice mitótico e índice de fases en los meristemos radiculares.
Material y métodos
Material biológico
Se utilizó Allium cepa L. “cebolla” para la ejecución de estudios de citotoxicidad (Grand, 1982; Becking et al., 1991; Fiskesjö, 1993), especie registrada con código 60877 en Herbarium Truxillense (HUT) de la Universidad Nacional de Trujillo.
Se usó el acetato cúprico Cu (CH3COO)2 como agente tóxico, la solución stock se preparó y las diferentes concentraciones a ser evaluadas fueron: 500 µg/l, 600 µg/l, 700 µg/l y 800 µg/l.
Bioensayo (Test Allium)
Consiste en exponer los bulbos de cebolla frente a diferentes concentraciones de cobre, a fin de determinar la citotoxicidad en células meristemáticas a nivel de raíz de Allium cepa L. (Díaz et al., 2004; De La Peña-Álvarez et al., 2012).
Método
Se seleccionó 25 bulbos de Allium cepa L., considerando su uniformidad en tamaño y semejantes características morfológicas. Luego cada bulbo de cebolla fue colocado en un recipiente con un contenido de 100 ml de agua destilada, teniendo el cuidado que se produzca el contacto del líquido con el disco germinativo. El periodo de crecimiento fue de 3 días, cuya longitud de las raicillas se encontraron entre de 2 a 3 cm, en el día 4 se colocaron los bulbos de A. cepa en contacto con las soluciones previamente preparadas a las concentraciones 500 µg/l, 600 µg/l, 700 µg/l y 800 µg/l. asegurándose que la zona radicular se encuentre en contacto con las soluciones durante 72 horas. Se procedió a coloración con orceína y HCL 1N (Tjio & Levan, 1950). Cada muestra fue observada con microscopio compuesto (Euromex, Olympus CX21), objetivo 40x y los recuentos de células meristemáticas con objetivo 100x.
Diseño experimental
En el diseño experimental se usó estímulo creciente a diferentes concentraciones de cobre para determinar el grado de toxicidad en Allim cepa L. (Cumpa-Yupton & Zavala-De La Cruz, 2013).
Determinación del índice mitótico e índice de fases
El porcentaje del índice mitótico se calculó mediante la siguiente fórmula según (Tjio & Levan, 1950).

Donde:
IM: Índice mitótico.
NCM: Número de células que están en mitosis.
NTC: Número total de células observadas.
El porcentaje del índice de fases se calculó mediante la siguiente fórmula según (Tjio & Levan, 1950).
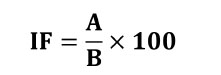
Donde:
IF: Índice de fases.
A: Número de las células de la fase que se encuentre.
B: Número total de células en mitosis.
Análisis estadístico
Para el análisis estadístico de la data se usó el programa estadístico InfoStat (versión 2008), la prueba de ANAVA y prueba de Tukey (Wayne, 2005).
Resultados y discusión
Tabla 1: Número de células meristemáticas en división de Allium cepa L. en los diferentes tratamientos con Cu2+ en condiciones de laboratorio
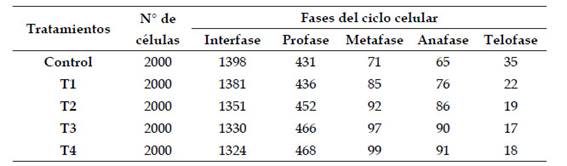
Nota: Tratamientos con cobre, según las concentraciones y tiempo de exposición
Control: (0 µg/l) de cobre, 36 horas.
T1: (500 µg/l) de cobre, 36 horas.
T2: (600 µg/l) de cobre, 36 horas.
T3: (700 µg/l) de cobre, 36 horas.
T4: (800 µg/l) de cobre, 36 horas.
Tabla 2: Índice mitótico e índice de fases de Allium cepa L. en los diferentes tratamientos con Cu2+ en condiciones de laboratorio
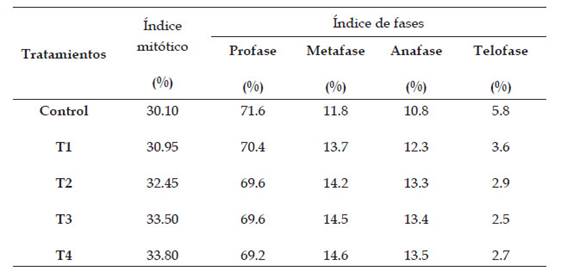
Tabla 3: Análisis estadístico de ANAVA y de comparación múltiple de medias por medio de la prueba de Tukey (p˂0.05) del índice profásico en células meristemáticas de Allium cepa L. “cebolla” sometidas a los tratamientos con Cu2+
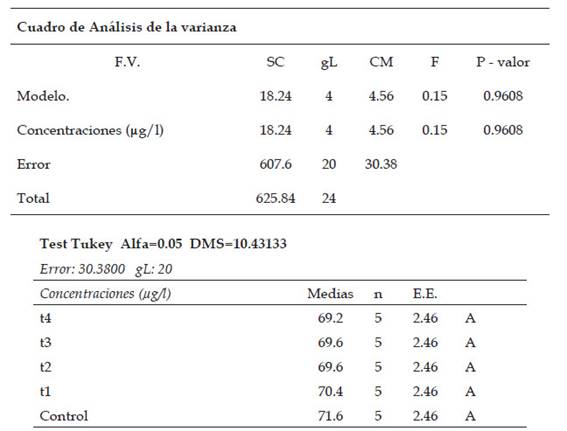
Tabla 4: Análisis estadístico de ANAVA y de comparación múltiple de medias por medio de la prueba de Tukey (p˂0.05) del índice metafásico en células meristemáticas de Allium cepa L. “cebolla” sometidas a los tratamientos con Cu2+
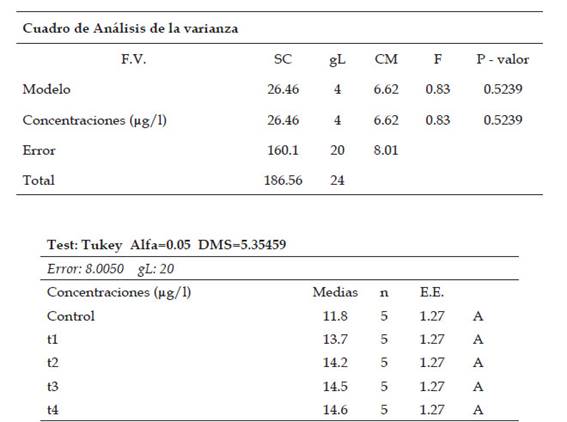
Tabla 5: Análisis estadístico de ANAVA y de comparación múltiple de medias por medio de la prueba de Tukey (p˂0.05) del índice anafásico en células meristemáticas de Allium cepa L. “cebolla” sometidas a los tratamientos con Cu2+
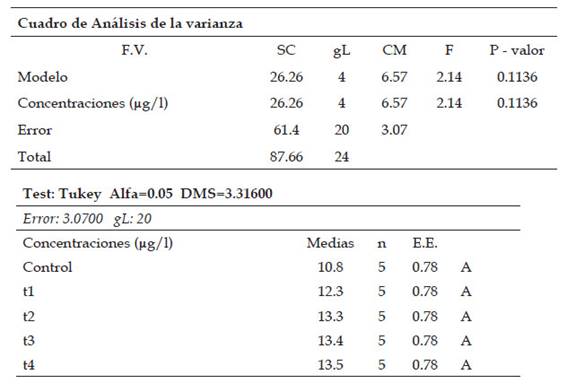
Tabla 6: Análisis estadístico de ANAVA y de comparación múltiple de medias por medio de la prueba de Tukey (p˂0.05) del índice telofásico en células meristemáticas de Allium cepa L. “cebolla” sometidas a los tratamientos con Cu2+
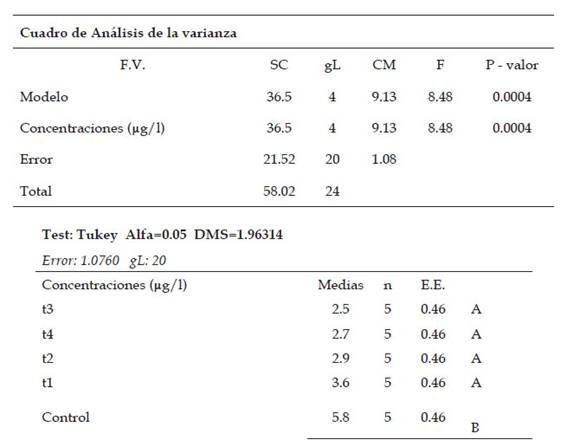
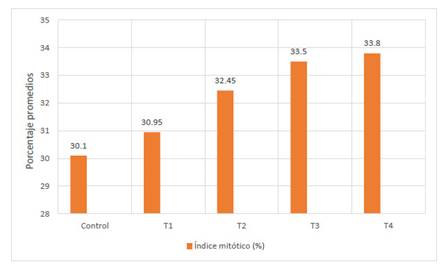
Fig. 1 Porcentaje promedio del índice mitótico en Allium cepa L. en los diferentes tratamientos con Cu+2 en condiciones de laboratorio
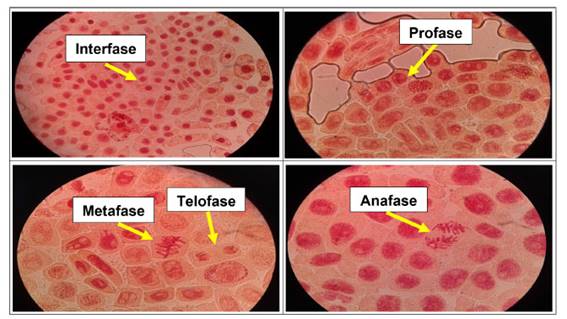
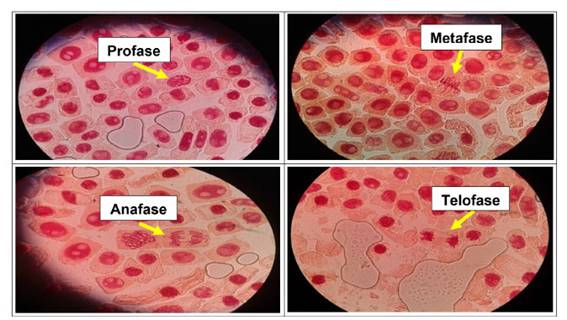
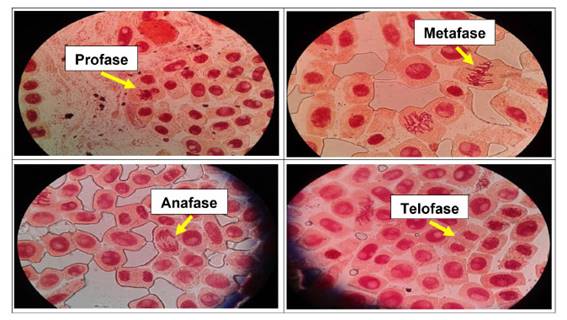
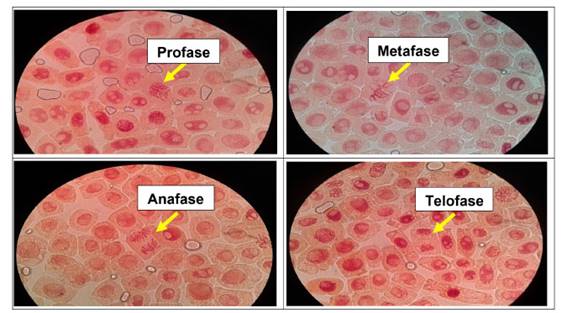
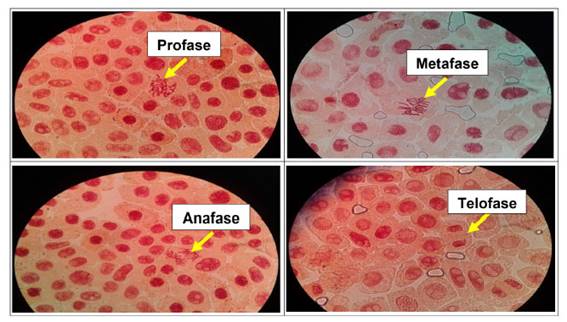
Se determinó anormalidades celulares en las concentraciones de 500 µg/l, 600 µg/l, 700 µg/l y 800 µg/l de Cu2+, a excepción del control. Las formas de anomalías encontradas fueron daños nucleares como rompimiento de cromosomas tanto en metafase como en anafase y telofase. En las concentraciones más altas (700 µg/l y 800 µg/l) se encontró una mayor cantidad de anomalías, así como daños nucleares, puentes anafásicos y telofásicos, sin embargo en las primeras dos concentraciones no existió daños en gran medida a nivel de las células meristemáticas.
En las figuras 7, 8, 9, 10, 11 y 12 se observaron daños en las diferentes concentraciones principalmente: metafase y anafase con puentes entre cromosomas, células alargadas tanto en profase como en telofase y rompimiento de los cromosomas en anafase y metafase.
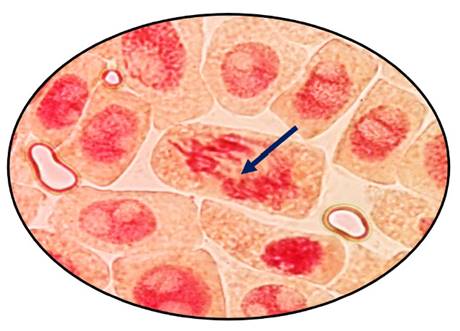
Fig. 7 Puente anafásico en anafase expuestas a la concentración de 700 µg/l de Cu+2 en Allium cepa L. “cebolla” a 100 x
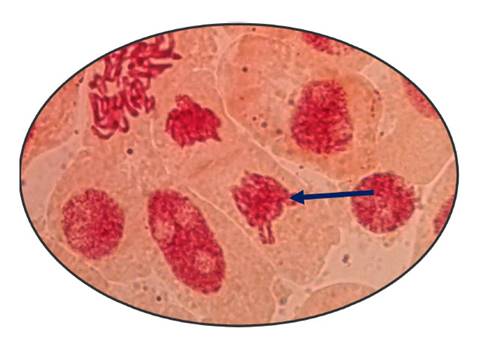
Fig. 8 Alargamiento de cromosomas en telofase expuestas a la concentración de 800 µg/l de Cu+2 en Allium cepa L. “cebolla” a 100 x
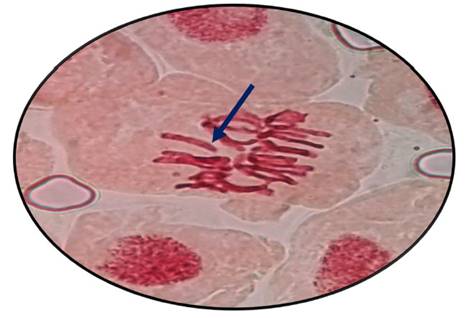
Fig. 9 Rompimiento de cromosomas en metafase expuestas a la concentración de 700 µg/l de Cu+2 en Allium cepa L. “cebolla” a 100 x
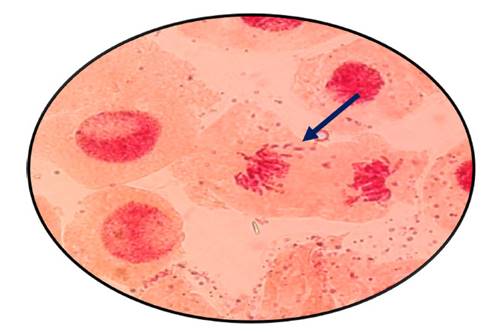
Fig. 10 Rompimiento de cromosomas en anafase expuestas a la concentración de 800 µg/l de Cu+2 en Allium cepa L. “cebolla” a 100 x
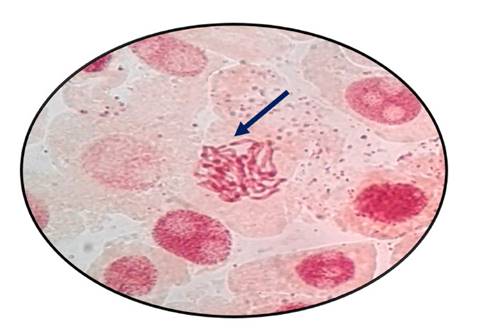
Fig. 11 Fragmentación de cromosomas en profase expuestas a la concentración de 800 µg/l de Cu+2 en Allium cepa L. “cebolla” a 100 x
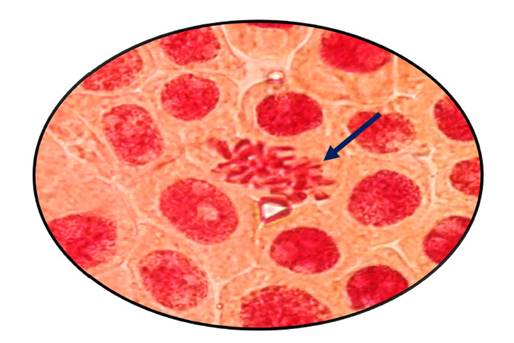
Fig. 12 Alargamiento de cromosomas en profase expuestas a la concentración de 800 µg/l de Cu+2 en Allium cepa L. “cebolla” a 100 x
Los tejidos meristemáticos se encuentran presentes en los ápices radiculares de Allium cepa L. “cebolla” es importante para el análisis de los ciclos celulares, debido a que, se desarrollan en un equilibrio constante; de esta forma al realizar el recuento de las células en cada fase mitótica, éstas son proporcionales a la duración de las mismas, esto es, cuando están en contacto con sustancias tóxicas, alteran su desarrollo normal e inclusive tiene la posibilidad de ocasionar daños a nivel cromosómico, provocando que la división celular pueda impedirse, retrasarse o provocar muerte celular (Díaz et al., 2004).
Las etapas del ciclo celular están controlados por los denominados complejos ciclinas-kinasas que dependen de ciclinas (CDK); las cuales son las encargadas de regular la transcripción y síntesis protéica que establecen el inicio del periodo celular y el paso siguiente a la G2, donde se producen sucesos involucrados a la condensación de los cromosomas, formación de husos cromosómicos y la desintegración de la envoltura nuclear como acontecimientos anteriores a la mitosis, que garantiza una correcta distribución del material cromosómico en las células hijas, por medio de la fluidez de etapas (Talledo, 1995).
Se determinó que existe variación en los ciclos celulares, en este caso un ligero crecimiento en el número de células en mitosis de las concentraciones de 500 µg/l, 600 µg/l, 700 µg/l y 800 µg/l de Cu2+ en opuesto al grupo control (Tabla 2). Los diversos estudios en Allium cepa L. evidenciaron que diversas sustancias de origen química u orgánica tienen la posibilidad de provocar un incremento o disminución del proceso mitótico durante la división celular en las células meristemáticas de raicillas de A. cepa. La variación del índice mitótico podría ser gracias al bloqueo del agente tóxico en la mitosis, incrementando o disminuyendo las células en separación, dado que el índice mitótico es una estimación de un valor cuantitativo en las fases de la mitosis en las células meristemáticas a nivel de las raicillas de la especie en estudio (Berrocal et al., 2013).
El índice mitótico (IM) es una medición de citotoxicidad que se puede realizar en cualquier ser vivo. El grado de citotoxicidad se establece cuando disminuye el número de fases del ciclo celular. Cuando la disminución del IM está por debajo del 22% en comparación con el control, entonces podría tener un efecto letal en los organismos, entretanto, si la disminución está por debajo del 50% en tal caso se tendría efectos negativos a largo plazo, a esto se llama valor límite de citotoxicidad (Prajitha & Thoppil, 2016).
Las alteraciones cromosómicas, se producen cuando existen cambios a nivel estructural y en el número de cromosomas, como el resultado de estar expuestos a agentes físicos o químicos (Ortiz et al., 2012; Hemanth et al., 2015). En el análisis llevado a cabo, las raicillas fueron sometidas a concentraciones de Cu2+ provocando diversas anomalías como puentes anafásicos y metafásicos y además se produjo la ruptura de los cromosomas a medida que se incrementó la concentración de cobre en estudio (Fig. 7, 8, 9, 10, 11 y 12).
El crecimiento significativo de los índices mitóticos en 30.10% (control: 0 µg/l de cobre, 36 horas), 30.95% (T1: 500 µg/l de cobre, 36 horas), 32.45% (T2: 600 µg/l de cobre, 36 horas), 33.50% (T3: 700 µg/l de cobre, 36 horas) y 33.80% (T4: 800 µg/l de cobre, 36 horas) con respecto al control demostraría la acción citotóxica de acetato cúprico en el ciclo celular de Allium cepa L. “cebolla”, se debe a la presencia de cobre, cuyo incremento de la concentración de cobre generaría la activación de la proteína quinasa, dicha proteína puede tener una participación en la señalización y fosforilación de proteínas por metales pesados (Yruela, 2005). Estudios realizados confirman la participación tiene la proteína quinasa en la señalización y en el tipo de relación que presenta con la reacción de oxidación-reducción en hojas y raíces en Arabidopsis thaliana sometidas al estrés por cobre, donde se concluyó que la proteína quinasa tiene una función vital en la transmisión de señales a nivel celular (Cuypers et al., 2013).
En la determinación del índice profásico no presentó diferencias significativas en la comparación de medias. El decrecimiento del índice profásico fue de 69,2 % (T4: 800 µg/l de cobre, 36 horas) con respecto al control, presentó un mayor porcentaje de citotoxicidad en comparación de los tratamientos y el control. La disminución del índice profásico a nivel de los tratamientos con Cu2+ inactiva la formación de los complejos CdK1-Ciclinas Ay B; lo cual daría como resultado la condensación del material genético y de la formación de los husos mitóticos dañados, en consecuencia, las células no podrían pasar de la profase a metafase. (Arnell et al., 2002; Hirano & Losada, 2001).
El índice metafásico no presentó diferencias significativas con respecto a los tratamientos y al control, con el porcentaje en incremento a partir de 11.8% (control: 0 µg/l de cobre) hasta 14.6% (T4: 800 µg/l de cobre), indicando un aumento en los daños a nivel celular en el proceso mitótico.
En el índice anafásico existió diferencias no significativas entre los tratamientos, presentaron que el valor se incrementa desde 10,8% (control: 0 µg/l de cobre) hasta 13,5% (T4: 800 µg/l de cobre), lo cual explica el desarrollo de células con anomalías en la etapa anafásica. Para el índice telofásico se encontró diferencias significativas entre los tratamientos (T1, T2, T3 y T4) a diferencia del control. Los valores fueron en incremento desde 2,5% (T3: 700 µg/l de cobre) hasta 5.8% (control: 0 µg/l de cobre), indicando un aumento en el número de anomalías cromosómicas entre los tratamientos.
Conclusiones
El efecto citotóxico de cobre en Allium cepa L. “cebolla” a diferentes concentraciones, se presentó en mayor porcentaje promedio en el tratamiento 4.
El índice mitótico en las células meristemáticas a nivel de las raicillas de Allium cepa L. “cebolla” a diferentes concentraciones de cobre fueron de 30,95% en tratamiento 1, 32,45% en tratamiento 2, 33,50% en tratamiento 3 y 33,80% en tratamiento 4.

