











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
La Fitosociología es una ciencia ecológica que trata de establecer el paralelismo entre unidades de vegetación y sus condiciones ambientales, permitiendo profundizar en el análisis y clasificación de los ecosistemas y, además, en la comparación ecológica, dinámica y geográfica con otras áreas disyuntas, resultados que no podemos obtener con estudios puramente fisionómicos (Moravec, 1992). La base de la metodología fitosociológica es el levantamiento de inventarios (plots) sobre fisiotopos o lugares geomorfológicos determinados (Schwabe & Kratochwil, 1994). Un inventario es un listado de especies acompañados de índices que, sobre todo, representan la estimación de la abundancia-dominancia de una planta en un punto. La repetición de un conjunto de plantas pertenecientes a distintas localizaciones geográficas constituye una asociación. Por tanto, la asociación es una imagen de la flora de un fisiotopo que presenta unas características dinámicas, corológicas e históricas. Las asociaciones se agrupan en alianzas, las alianzas en órdenes, y los órdenes en clases fitosociológicas (Braun-Blanquet, 1932, Géhu & Rivas-Martínez, 1981, Dierschke, 1994, Rivas-Martínez, 2007, Dengler et al., 2008) de tal forma que podemos construir un ordenamiento jerárquico donde cada una de estas unidades tiene una distribución geográfica. Este orden jerárquico constituye el esquema sintaxonómico de la vegetación de un territorio, que es la Ɛ- diversidad, que expresa el número de comunidades vegetales de una región (Hecht et al., 1977, Halffter et al., 2001).
Hace ya años que publicamos un primer esquema sintaxonómico global de la Región del Caribe y América del Sur, desde el nivel de clase hasta el de alianza (Galán de Mera, 2005), basándonos en las descripciones de otros autores y en las nuestras propias, con trabajos que comprendían desde la vegetación de Cuba (Borhidi, 1996) a la de la Patagonia (Boelcke et al., 1985). Recientemente publicamos el primer esquema sintaxonómico del Perú (Galán de Mera et al., 2021), donde se recogen desde las unidades fitosociológicas de la costa hasta algunos de los bosques húmedos de la Amazonía. Sin embargo, en un país mega- diverso, siempre van a aparecer nuevas unidades. En este trabajo añadimos algunas correspondientes a inventarios nuevos y otros que quedaron sin ver la luz.
Material y métodos
En este trabajo presentamos tablas, descripciones y arreglos nomenclaturales correspondientes a unidades de vegetación de los departamentos de Arequipa, Cusco y Puno. Los plots de las tablas fueron levantados según la metodología fitosociológica de Braun-Blanquet (1932), con la siguiente escala de abundancia- dominancia: + individuos escasos, de débil cobertura (< 1%), 1- individuos bastante abundantes pero de débil cobertura (1- 5%), 2- individuos que cubren 5-25% de la superficie, 3- individuos que cubren 25-50% de la superficie, 4- individuos que cubren 50-75% de la superficie, y 5- individuos que cubren > 75% de la superficie. Para ello se tiene en cuenta que la abundancia- dominancia es evaluada estrato por estrato. Las coordenadas de la localidad de cada inventario se expresan en “Universal Transversal de Mercator”.
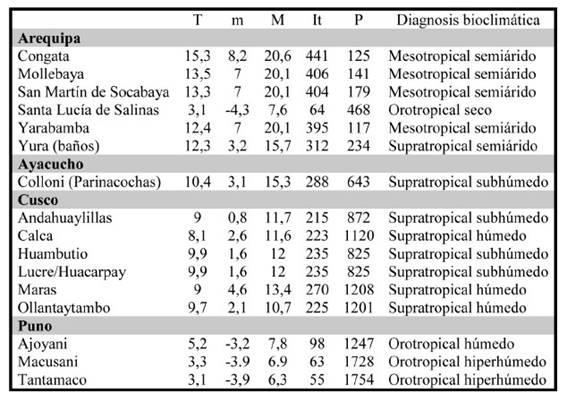
Las asociaciones se correlacionan con pisos bioclimáticos según la metodología de Rivas-Martínez (Rivas-Martínez et al., 1999), desarrollados y cartografiados para el Perú por Galán de Mera et al. (2017). Los pisos bioclimáticos son cinturas altitudinales y latitudinales de vegetación basadas en intervalos del índice de termicidad (It), compuesto por valores de temperatura en grados Celsius [It = (T+m +M) 10; T= temperatura media anual, m= temperatura media de las mínimas del mes más frío, M= temperatura media de las máximas del mes más frío] y bioindicadores (plantas características de cada piso). A su vez, los pisos bioclimáticos se matizan con intervalos de precipitaciones en mm. En el Perú reconocemos 6 pisos bioclimáticos [infratropical (It > 690), termotropical (It = 490 a 690), mesotropical (It = 320 a 490), supratropical (It = 160 a 320), orotropical (It = 50 a 160) y criorotropical (It = < 50)] y 9 intervalos de precipitaciones [ultrahiperárido (P < 5), hiperárido (5 a 30), árido (31 a 100), semiárido (101 a 300), seco (301 a 500), subhúmedo (501 a 900), húmedo (901 a 1500), hiperhúmedo (1501 a 2500) y ultrahiperhúmedo (> 2500). En la tabla 1 se muestran los valores de temperaturas y precipitaciones, tomados de las bases de datos Chelsa (Karger et al., 2017) y Climate-Data (Schwarz, 2022), índice de termicidad (It) y diagnosis en las estaciones meteorológicas más próximas a los plots levantados y las observaciones realizadas.
Tabla 1 Valores de temperaturas (ºC) y precipitaciones (mm), índice de termicidad (It) y diagnosis bioclimática de las estaciones meteorológicas más próximas a los plots levantados y puntos con observaciones.
Las características geológicas de los territorios han sido tomadas del IGM (1975) y del Atlas Digital del Perú (2005).
La nomenclatura de las unidades fitoso- ciológicas sigue el Código Internacional de Nomenclatura Fitosociológica (Theurillat et al., 2021), y la de unidades biogeográfi- cas a Galán de Mera et al. (2021), que a su vez se fundamentan en Rivas-Martínez et al. (2011).
Para separar las diferentes unidades de vegetación de los saladares andinos y costeros hemos realizado un Análisis de Componentes Principales (ACP), con el programa PAST 4.07b (Hammer, 1999- 2021), usando inventarios de Bolivia (Navarro, 1993) y del Perú (Gutte & Müller, 1985; Galán de Mera et al., 2009, 2011). El desarrollo de un biplot con vectores nos da a conocer la tendencia de presencia de especies en los distintos grupos.
Resultados y discusión
Los matorrales del valle de Oropesa (Cusco)
Berberido bolivianae-Flourensietum polycephalae Galán, Linares, Montoya & Vicente ass. nov.
Tabla 2. holotypus: plot 4; Fig. 1
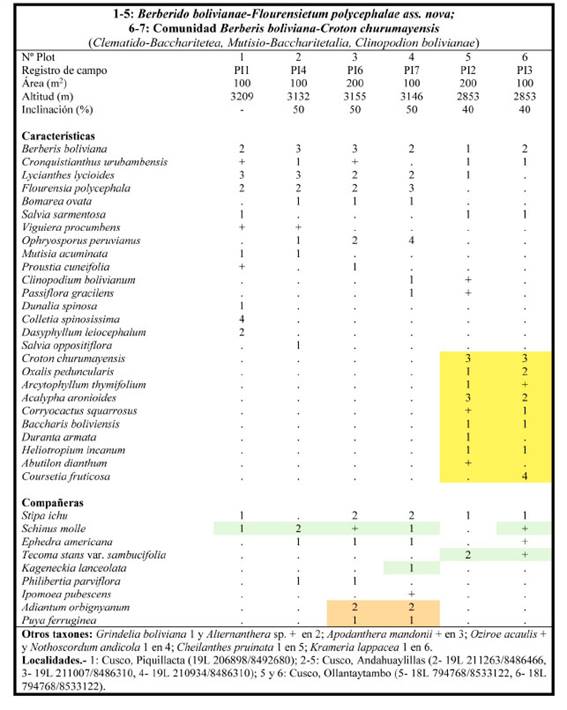

Fig. 1 Algarrobales y matorrales junto a las ruinas de Piquillacta (Huacarpay, Cusco). A- Schino mollis-Prosopidetum andicolae, B- Berberido bolivianae-Flourensietum polycephalae.
Características de asociación: Berberis boliviana Lechl., Corryocactus squarrosus (Vaupel) Hutchison, Cronquistianthus urubambensis (B.L.Rob.) R.M.King & H.Rob., Croton churumayensis Croizat, Flourensia polycephala M.O. Dillon, Heliotropium incanum Ruiz & Pav. y Salvia sarmentosa Epling.
Matorrales y arbustos con un dosel de unos 1,80-2 m de altura, que se encuentran generalmente en pendientes de fuerte inclinación, sobre suelos muy arcillosos, ricos en bases, de origen jurásico y cretácico, dentro del bioclima supratropical subhúmedo en el sector biogeográfico Cusco.
Es una asociación rica en endemismos como Coursetia fruticosa (Cav.) J. F. Macbr. (endemismo de las provincias Puneña Mesofítica y Yungueña, 1500-3000 m), Corryocactus squarrosus (endemismo de provincia Puneña Mesofítica, 2850-3000 m), Cronquistianthus urubambensis (endemismo de la provincia Puneña Mesofítica, 2800- 3300 m), Croton churumayensis (endemismo del sector de Cusco, 2000-4000 m), Duranta armata Moldenke (endemismo de las provincias Puneña Mesofítica y Yungueña, 1400-3800 m), Flourensia polycephala (endemismo del sector de Cusco, entre 2800-4000 m), Salvia oppositiflora Ruiz & Pav. (endemismo peruano, 700-3900 m) y Salvia sarmentosa (sectores de Cusco y Abancay, 2500-4000 m).
Estos matorrales pueden presentar algunas variantes: Los inventarios 5 y 6 contienen una serie de elementos (Tabla 2, color anaranjado) que se desarrollan a menor altitud en el piso bioclimático mesotropical. Coursetia fruticosa, Croton churumayensis y Duranta armata alcanzan la ceja de montaña, por lo que podemos hablar de una variante más térmica. La presencia en varios plots con pequeños árboles (Kageneckia lanceolata Ruiz & Pav., Schinus molle L. y Tecoma stans var. sambucifolia (Kunth) J.R.I.Wood) (Tabla 2, color verde) indican la acumulación de agua en el subsuelo por la presencia de arcillas, característica ecológica que también presentan la asociación Cynancho tarmensis-Tecometum sambucifoliae (Galán de Mera, 1996) y la comunidad de Lomanthus velardei (Cabrera) B. Nord. & Pelser (S. velardei Cabrera) y Tecoma stans var. sambucifolia (Galán de Mera et al., 2004), ambas del centro del Perú. Adiantum orbignyanum Mett. y Puya ferruginea (Ruiz & Pav.) L. B. Sm. indican medios pedregosos (Tabla 2, color ocre), situación que también comentan Seibert & Menhofer (1991) para
A. orbignyanum, que lo incluyen como una característica de este tipo de matorrales, aunque en realidad forma una comunidad con caracteres ecológicos diferenciados propios de la clase Adiantetea capilli-veneris Br.-Bl., Roussine & Nègre 1952.
En nuestra tabla y en nuestro territorio aparece la especie oriental Clinopodium bolivianum (Benth.) Kuntze (Satureja boliviana (Benth.) Briq.), directriz de la alianza Clinopodion bolivianae Seibert 1993 nom. mut. nov. (Seibert, 1993). Colletia spinosissima J. F.Gmel., Dunalia spinosa (Meyen) Dammer, Mutisia acuminata Ruiz & Pav., Ophryosporus peruvianus (J. F. Gmel.) R. M. King & H. Rob., Proustia cuneifolia D. Don y Viguiera procumbens (Pers.) S. F. Blake permiten la inclusión de Clinopodion bolivianae en el orden Mutisio acuminatae- Baccharitetalia latifoliae y en la clase Clematido peruvianae-Baccharitetea latifoliae (Galán de Mera et al., 2015). De esta forma el orden está constituido en el Perú por tres alianzas: Mutisio acuminatae-Ophryosporion peruviani Galán & Cáceres in Galán, Rosa & Cáceres 2002, occidental del centro y sur del Perú, Otholobio munyensis-Rubion robusti Galán, Sánchez, Montoya, Linares, Campos & Vicente 2015, de los Andes del norte del Perú, Ecuador y ceja de selva oriental del país, y Clinopodion bolivianae, de los Andes orientales de Perú y Bolivia.
Los algarrobales de los valles de Oropesa y Urubamba (Cusco)
Schino mollis-Prosopidetum andicolae Galán, Linares, Montoya & Vicente ass. nov.
Tabla 3; holotypus: plot 6; Fig. 1
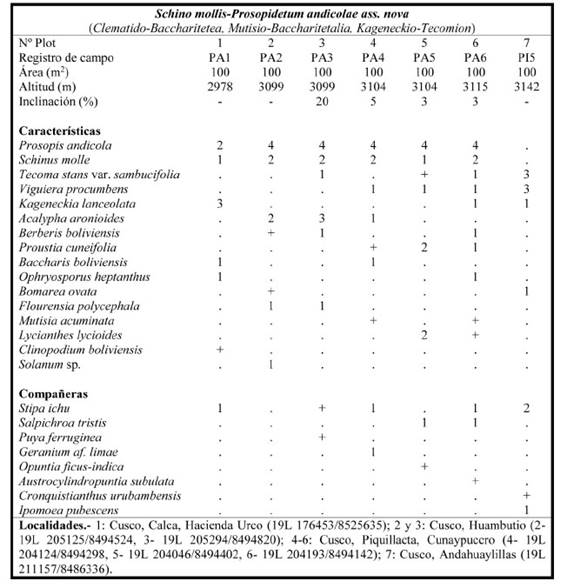
Características de asociación: Kageneckia lanceolata, Prosopis andicola (Burkart) A.Galán, E.Linares, J.Montoya & Vicente Orell., Schinus molle y Tecoma stans var. sambucifolia.
Bosques con dosel de entre 4-6 m de altura, cuya especie dominante es Prosopis andicola (“thacco”), que se encuentra acompañada de otros árboles como Schinus molle, Tecoma stans var. sambucifolia y Kageneckia lanceolata que se asientan sobre los suelos calizos muy arcillosos, con escasa pendiente del sector biogeográfico de Cusco, en el piso bioclimático supratropical subhúmedo. Normalmente se sitúan en las zonas basales de las lomadas con poco relieve donde hay acumulación de humedad, de ahí la constancia de Schinus molle y Tecoma stans var. sambucifolia, contactando con las especies de la asociación anterior.
Como ya apuntaba Venero et al. (2017) estos bosques están muy alterados y P. andicola ha desaparecido de muchas localidades y en otras solo quedan ejemplares aislados. En situaciones como las del inventario 7, donde figuran los principales elementos del bosque, P. andicola tal vez haya desaparecido. Es por ello que solo hemos podido levantar inventarios en las cercanías de las ruinas incaicas de Urco (Calca), y en las proximidades de las ruinas Wari de Piquillacta. Al parecer hubo una deforestación en los valles de Oropesa y Urubamba desde hace siglos, si nos atenemos al dato de que los Incas esquilmaron los bosques de la región de Cusco en favor de las construcciones de los techos de los edificios de la ciudad, a pesar de que tenían funcionarios responsables del cuidado de los bosques (“mallki kamalloc”) (Arguto Calvo, 1988).
Las asociaciones basófilas con bajo dosel arbóreo Cynancho tarmensis-Tecometum sambucifoliae, del centro del Perú (Galán de Mera, 1996), y Schino mollis-Prosopidetum andicolae del sur las integramos dentro de la nueva alianza de la provincia biogeográfica Puneña Mesofítica Kageneckio lanceolatae- Tecomion sambucifoliae Galán, Linares & Montoya all. nov. (holotypus: Schino mollis- Prosopidetum andicolae; características de la alianza: Cynanchum tarmense Schltr., Helogyne virgata B.L.Rob., Kageneckia lanceolata, Prosopis andicola, Schinus molle (territorial), Lomanthus tovarii (Cabrera) B.Nord. & Pelser (S. tovarii Cabrera) y Tecoma stans var. sambucifolia). Según la composición florística de la tabla 3, esta nueva alianza se incluye en el orden Mutisio acuminatae-Baccharitetalia latifoliae y en la clase Clematido peruvianae-Baccharitetea latifoliae.
Los algarrobales de Arequipa
Schino mollis-Prosopidetum calderensis Galán, Linares, Montoya & Vicente ass. nov.
Tabla 4; holotypus: plot 2; Fig. 2
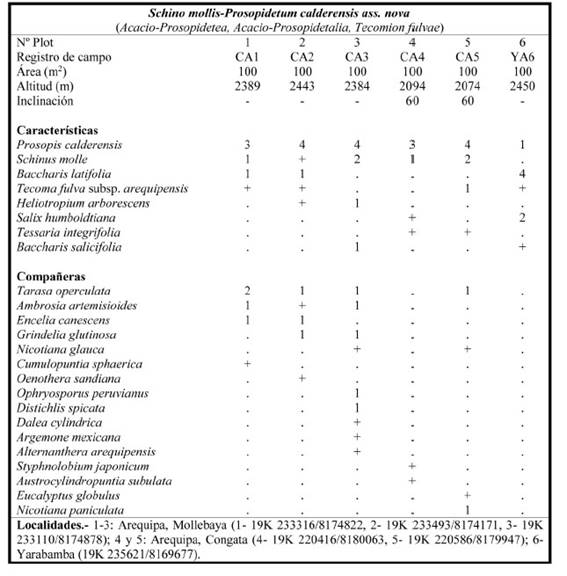

Fig. 2 Algarrobales en el departamento de Arequipa. A- Schino mollis-Prosopidetum calderensis en Mollebaya, B- Aspecto invernal de Schino mollis-Prosopidetum calderensis en una quebrada de Mollebaya, que sufre alteración por una invasión humana.
Características de asociación: Baccharis latifolia Pers., B. salicifolia (Ruiz & Pav.) Pers., Heliotropium arborescens L., Prosopis calderensis A.Galán, E.Linares, J.Montoya & Vicente Orell., Salix humboldtiana Willd., Schinus molle, Tecoma fulva subsp. arequipensis (Sprague) J.R.I.Wood y Tessaria integrifolia Ruiz & Pav.
Prosopis calderensis (“yaro”) fue descrita separando las poblaciones de los valles del interior de Cusco de las del Batolito de la Caldera en Arequipa (Galán de Mera et al., 2019), que evidentemente tienen una ecología y biogeografía diferentes, aunque ambas tienen preferencias por los suelos ricos en bases.
Esta nueva asociación representa los bosques de freatófitos de las quebradas secas, pedregosas y arenosas, con materiales jurásicos y cretácicos de calizas y afloramientos de yesos, bajo el bioclima termo-mesotropical árido e incluso hiperárido, del sector biogeográfico de Arequipa. La especie dominante es Prosopis calderensis que se muestra en poblaciones con un dosel de hasta 4 m de altura, que en ocasiones hemos visto en suelos costrosos poligonales (Yarabamba) formando comunidades de gran similitud con las de Prosopis tamarugo Phil. en Iquique (Chile).
Las comunidades de freatófitos de las quebradas occidentales del sur del Perú se incluyen en la alianza Tecomion fulvae, que se extiende desde Ica a Arica (Chile) y tal vez más hacia el Sur (Wood, 2008). Recientemente, Chicalla-Ríos (2021) mostraba la extensión de las características de esta asociación hasta la cuenca del río Tambo (Moquegua), en las cercanías de Coalaque, describiendo una comunidad de Tecoma fulva subsp. arequipensis y Prosopis calderensis, que sin duda son fragmentos de esta nueva asociación en sus altitudes más elevadas, hacia los 2500 m, tal como ocurre igualmente en las proximidades de Polobaya (Arequipa). Este autor incluye a esta vegetación en la clase de comunidades de cactáceas Opuntietea sphaericae Galán & Vicente 1996 sin advertir que tanto Tecoma fulva subsp. arequipensis como Prosopis calderensis son freatófitos que quedan englobados en la vegetación azonal del sur del Perú, mientras que Opuntietea sphaericae reúne sintáxones de vegetación zonal, con una ecofisiología muy diferente. Por otra parte, este mismo autor incluye entre sus especies representativas a un pul de plantas que, como Eragrostis nigricans (Kunth) Steud., Galium corymbosum Ruiz & Pav. o Pectocarya linearis (Ruiz & Pav.) DC. (P. lateriflora (Lam.) DC.), pertenecen a la clase Crassuletea connatae Galán 1999 de vegetación terofítica pastoreada (Galán de Mera et al., 2012).
Los saladares meso-supratropicales del sur del Perú
Hasta la actualidad, solo conocíamos comunidades vegetales de suelos salinos correspondientes a las clases Distichlio humilis-Anthobryetea triandri Navarro 1993, donde intervienen Salicornia pulvinata R. E. Fr. y Frankenia triandra Remy (Anthobryum triandrum (Remy) Surgis) en el piso orotropical semiárido-seco en Salinas (Arequipa) (Galán de Mera et al., 2011), y a la clase litoral Batido-Salicornietea ambiguae Borhidi 1996, con asociaciones con Salicornia neei Lag. y Distichlis spicata (L.) Greene (Galán de Mera et al., 2009).
Sin embargo, existe vegetación meso- supratropical halófila que se extiende entre las salinas de Yura (Arequipa), la laguna de Parinacochas (Ayacucho), el saladar de Huacarpay (Cusco) y las salinas de Maras (Cusco), con una composición florística diferenciada, donde Distichlis spicata sigue teniendo importancia al igual que en la costa.
En el análisis de componentes principales (ACP) de la figura 3 podemos distinguir cuatro grupos: A- son las asociaciones orotropicales de Bolivia (Navarro, 1993), donde Distichlis humilis Phil., Frankenia triandra y Salicornia pulvinata son las especies más preponderantes; B- las asociaciones de la costa del centro y sur del Perú (Galán de Mera et al., 2009), con Salicornia neei, Cressa truxillensis Kunth y Sporobolus virginicus (L.) Kunth; C1- la asociación con Salicornia cuscoensis Gutte & G. K. Müll. ex Freitag, M. Á. Alonso & M. B. Crespo de Gutte & Müller (1985) en los saladares de Cusco, y C2- los plots de la asociación con Distichlis spicata de Gutte & Müller (1985) en Huacarpay (Cusco) y de Galán de Mera et al. (2011) en Yura (Arequipa). Los plots de Cusco y Arequipa se encuentra bien separados del resto puesto que intervienen Salicornia cuscoensis o Distichlis spicata con otras plantas, como Hymenoxys robusta (Rusby) K. F. Parker, lo que nos permite suponer unidades fitosociológicas diferentes propias de los pisos meso- y supratropical.

Fig. 3 Análisis de Componentes Principales (ACP) con diferentes agrupaciones de plots levantados sobre suelos salinos. A- Plots de Bolivia (nt; Navarro, 1993) con Distichlis humilis, Frankenia triandra y Salicornia pulvinata, B- Plots de la costa peruana (ca; Galán de Mera et al., 2009) con Salicornia neei, Cressa truxillensis y Sporobolus virginicus, C1- Plots de Cusco (gm; Gutte & Müller, 1985) con Salicornia cuscoensis, C2- Plots de Cusco (gm; Gutte & Müller, 1985) y de Yura (li; Galán de Mera et al., 2011) con Distichlis spicata y otras especies no litorales. Los vectores indican las especies preponderantes en cada grupo. Las componentes son un gradiente de aparición o ausencia de determinadas especies según las áreas geográficas.
Salicornietum cuscoensis Gutte & Müller 1985
Vegetación almohadillada suculenta que se asienta en los suelos salinos que se encharcan intermitentemente, de color marrón oscuro, muy arcilloso, y una capa de sal en la superficie, bajo un bioclima supratropical subhúmedo, al menos en los sectores biogeográficos de Cusco y Xauxa, pues Salicornia cuscoensis fue descrita con material de Huacarpay (Cusco) y Parinacochas (Ayacucho) (Alonso et al., 2017) (Fig. 4).

Fig. 4 Detalle de Salicornia cuscoensis (A) y de la asociación Salicornietum cuscoensis (B) en la laguna de Huacarpay (Cusco).
Gutte & Müller (1985) al describir la asociación, lo hicieron sobre el nomen nudum de Salicornia cuscoensis, con lo cual el nombre de la asociación quedaba inválido (Art. 3g y 3l), llevándonos a la descripción de Sarcocornietum andinae (Galán de Mera et al., 2021), pues Alonso & Crespo (2008) llevaron a Salicornia andina Phil. (Sarcocornia andina (Phil.) Freitag, M. A. Alonso & M. B. Crespo) hasta la laguna de Parinacochas (Ayacucho), interpretando a S. cuscoensis como una variante fenotípica de esta. Tras la descripción de S. cuscoensis (Alonso et al., 2017), el nombre Salicornietum cuscoensis queda validado (Art. 2), S. andina se reparte por las altitudes medias Argentina, Bolivia y Chile, y S. pulvinata vive por encima de los 3500 m en Argentina, Bolivia, Chile y Perú (Alonso & Crespo, 2008; Galán de Mera et al., 2011).
Hymenoxyo robustae-Distichlietum spicatae Galán, Linares, Montoya & Vicente nom. nov.
Vegetación dominada por Distichlis spicata que se asienta sobre suelos más elevados y secos que la asociación anterior. Cuando el suelo comienza a encharcarse, penetra en la asociación Schoenoplectus pungens var. badius (C. Presl) S. G. Sm. (Scirpus olneyi A. Gray) que se va haciendo dominante a medida que aumenta el nivel de inundación (Fig. 5).

Fig. 5 Disposición de la vegetación en la laguna de Huacarpay (Cusco). A- Salicornietum cuscoensis, B- Hymenoxyo robustae-Distichlietum spicatae, C- Schoenoplectetum badii, D- Schoenoplectetum tatorae.
En la laguna de Huarcapay no existe Distichlis humilis tal como indicaban Gutte & Müller (1985), sino D. spicata, puesto que las plantas tienen una altura de más de 10 cm presentando numerosas espiguillas (Tovar, 1993). Por tanto, es necesario corregir la asociación Distichlietum humilis (Art. 43), pero entonces interfiere con el Distichlietum spicatae Ciferri 1936 de la Española y Cuba (Borhidi et al., 1979) y Distichlietum spicatae López Guillén & Rivas Goday 1952 de Lima, llevándonos a crear un nombre nuevo (Art. 43). En la tabla 2 de Gutte & Müller (1985) se cita a Hymenoxys anthemoides Cass. (H. haenkeana DC.), que debería considerarse en el nombre de la asociación si no fuese porque no es una especie peruana (Brako & Zarucchi, 1993) confundida con H. robusta, que también se encuentra en otras áreas salinas del valle de Oropesa y Urubamba (ver el pliego US 1340756).
La presencia de especies halófilas en los pisos meso- y supratropical de las áreas internas del Perú, y la ausencia de elementos de la clase Distichlio humilis- Anthobryetea triandri, nos permiten describir la alianza Salicornio cuscoensis-Distichlion spicatae Galán, Linares, Montoya & Vicente all. nov. (holotypus: Hymenoxyo robustae-Distichlietum spicatae; especies características: Distichlis spicata, Hymenoxys robusta, Spergularia marina (L.) Besser, Salicornia cuscoensis y Triglochin striata Ruiz & Pav.), el orden Hymenoxyo robustae- Distichlietalia spicatae Galán, Linares, Montoya & Vicente ord. nov. (holotypus: Salicornio cuscoensis-Distichlion spicatae; con las mismas especies características que la alianza), y la clase Hymenoxyo robustae- Distichlietea spicatae Galán, Linares, Montoya & Vicente cl. nov. (holotypus: Hymenoxyo robustae-Distichlietalia spicatae; con las mismas especies características). En el mapa de la figura 6 se representan las localidades con plots de las tres clases fitosociológicas de los saladares del Perú (Batido-Salicornietea ambiguae y Hymenoxyo robustae-Distichlietea spicatae) e Bolivia (Distichlio humilis-Anthobryetea triandri).
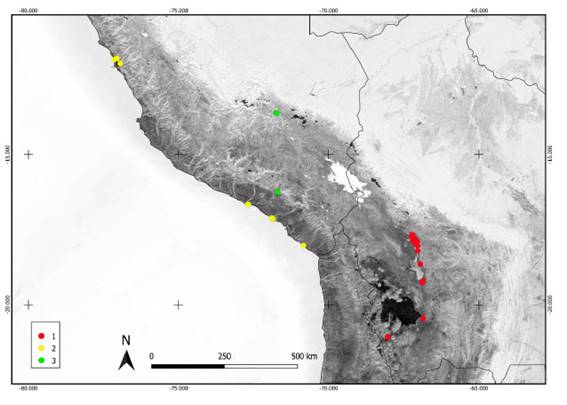
Fig. 6 Localización de los plots correspondientes a las clases fitosociológicas de saladares del Perú y Bolivia. 1. Distichlio humilis-Anthobryetea triandri, 2. Batido-Salicornietea ambiguae, Hymenoxyo robustae-Distichlietea spicatae.
Schoenoplectetum badii Galán, Linares, Montoya & Vicente ass. nov.
Características de asociación:
Schoenopletus pungens var. badius.
Plots: 1: Schoenoplectus pungens var. badius 5, Schoenoplectus tatora +, Distichlis spicata 1 (Cusco, Huacarpay, 19L 203922/8493463, 3079 m, 100 m2, profundidad 50 cm; holotypus); 2- Schoenoplectus pungens var. badius 4, Distichlis spicata +, Pennisetum clandestinum 1 (Arequipa, Yura, 19K 211640/8201956, 2501 m, 100 m2, profundidad 20 cm).
Asociación andina de juncales de pequeña talla (20-30 cm), de lagunas salobres que forman una banda interior más encharcada respecto a las comunidades de Distichlis spicata. Al ganar en profundidad da paso a la asociación Schoenoplectetum tatorae Galán 1995, nom. mut. Galán, Campos, Linares, Montoya, Torres & Vicente 2021 (Fig. 5). La comunidad de Nitella sp. y Schoenoplectus americanus (Pers.) Volkart mencionada por Montesinos- Tubée (2012) en San Martín de Socabaya (Arequipa) corresponde a esta asociación.
Esta nueva asociación está basada en Schoenoplectus pungens var. badius, que es el taxon que cita Smith (1995), cuyo material tipo procede de los Andes peruanos, y su composición florística con S. tatora, nos lleva a incluirla en la alianza Schoenoplection tatorae Galán, Campos, Linares, Montoya, Torres & Vicente 2021 del orden Schoenoplectetalia olneyi-americani Galán, Linares, Campos & Vicente 2009 y la clase neotropical Xyrido carolinianae-Typhetea domingensis O. Bolòs, Cervi & Hatschbach 1991.
Vegetación basófila en Puno
Poo candamoanae-Festucetum humilioris Galán, Linares, Montoya & Vicente ass. nova
Tabla 5; holotypus: plot 1; Fig. 7
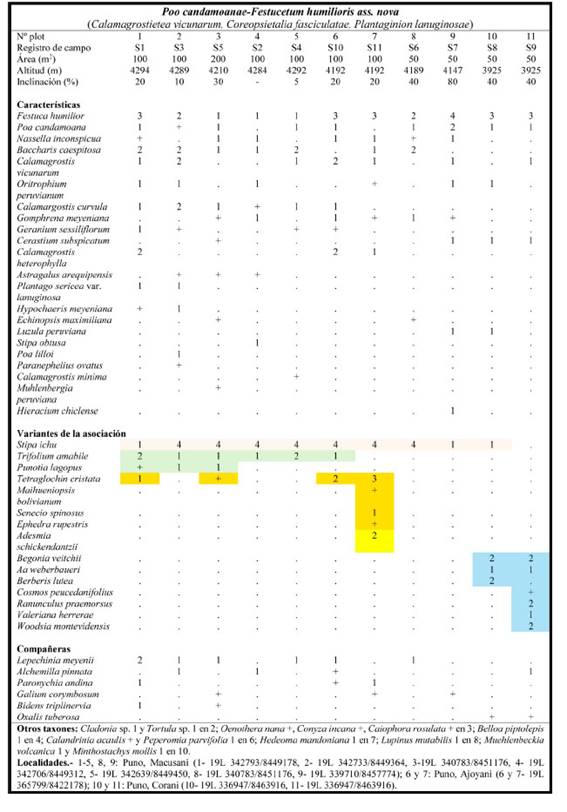

Fig. 7 Dos aspectos de la asociación Poo candamoanae-Festucetum humilioris: A- Stipa ichu junto a cultivos de papa y oca, B- Punotia lagopus en el seno de la asociación.
Características de asociación: Aa weberbaueri Schltr., Festuca humilior Nees & Meyen, Nassella inconspicua (J.Presl) Barkworth, Oritrophium peruvianum (Lam.) Cuatrec., Plantago sericea var. lanuginosa Griseb., Poa candamoana Pilg. y Punotia lagopus (K.Schum.) D.R.Hunt.
Pajonales supra- orotropicales húmedos-hiperhúmedos que se asientan sobre suelos profundos, pedregosos en superficie aunque muy arcillosos, calcáreos derivados de calizas cretácicas orientales, del sector biogeográfico del Allincapac.
Al igual que en otras áreas del Perú, los pajonales muestran una cierta variabilidad dependiendo del uso del territorio. La presencia de Stipa ichu (Ruiz & Pav.) Kunth (Tab. 5, color rosa) indica medios que han sido alterados por cultivos y por el pisoteo de la ganadería de camélidos (Fig. 7A), aspecto del dinamismo de la vegetación que ya fue observado tanto en el sur (Galán de Mera et al., 2003) como en el centro del Perú (Galán de Mera et al., 2004) y Bolivia (Seibert & Menhofer, 1992).
Trifolium amabile Kunth y Punotia lagopus (Tab. 5, color verde) son propias de un pastoreo más reciente, pues las alpacas remueven los tallos de P. lagopus (Fig. 7B) para obtener agua, y así la dispersan (comunicación de una pastora). La presencia de Tetraglochin cristata (Britton) Rothm. está indicando un aumento de la erosión del suelo, pero junto a otras plantas, como Ephedra rupestris Benth., Maihueniopsis bolivianum (Salm-Dyck) R. Kiesling y Senecio spinosus DC. (Tab. 5, color anaranjado) evoca la influencia de la alianza Azorello- Festucion orthophyllae (Galán de Mera et al., 2003) en áreas más secas y occidentales, al sur del paso de Oquepuño, donde además se integra Adesmia schickendantzii Griseb. (Tab. 5, color amarillo), en el extremo de su distribución desde Argentina y Bolivia (Linares Perea et al., 2015). Los plots 10 y 11 (Tab. 5, color azul) incluyen plantas fisurícolas que crecen a menor altitud, en el piso supratropical; son plots con una fuerte inclinación donde los pajonales alternan con las paredes de andenes muy antiguos con viejos cultivos de oca.
Debido a la composición florística de la tabla, incluimos esta nueva asociación en la alianza Plantaginion lanuginosae Gutte 1986 nom. mut. Galán, Campos, Linares, Montoya, Torres & Vicente 2021, y en el orden Coreopsietalia fasciculatae Galán, Campos, Linares, Montoya, Torres & Vicente 2021, que reúne a pajonales sobre substratos básicos (Gutte, 1986) dentro de la clase Calamagrostietea vicunarum Rivas- Martínez & Tovar 1982.
Woodsio montevidensis-Begonietum veitchii Galán, Linares, Montoya & Vicente ass. nova
Tabla 6; holotypus: plot 1; Fig. 8
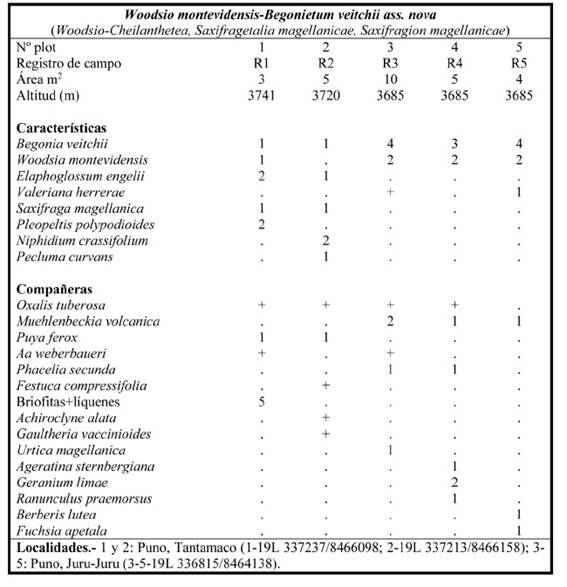

Fig. 8 Aspecto de la asociación Woodsio montevidensis-Begonietum veitchii en los andenes preincaicos de la ciudadela de Juru-Juru (Puno).
Características de asociación: Begonia veitchii Hook.f., Elaphoglossum engelii Christ, Niphidium crassifolium (L.) Lellinger, Pecluma curvans (Mett.) M.G.Price, Pleopeltis polypodioides (L.) E.G.Andrews & Windham, Saxifraga magellanica Poir., Valeriana herrerae Killip y Woodsia montevidensis Hieron..
Vegetación fisurícola basófila, supra- orotropical húmeda-hiperhúmeda oriental de los sectores biogeográficos Cusco y Allincapac, siguiendo la distribución de Begonia veitchii, que se adentra en Bolivia (Tropicos, 2022). Es una asociación propia de roquedos verticales y taludes (Fig. 8), pero también se pone de manifiesto en las paredes de los Andenes preincaicos del valle de San Gabán, donde abunda Oxalis tuberosa Molina (oca) que se encuentra por doquier escapada de cultivo.
Incluimos esta asociación en la alianza Saxifragion magellanicae Galán & Cáceres in Galán, Rosa & Cáceres 2002, junto a las asociaciones Valeriano thalictrioidis-Saxifragetum magellanicae Galán & Cáceres in Galán, Rosa & Cáceres 2002, del centro del Perú (sector de Xauxa) y Saxifrago magellanicae-Leucherietum daucifoliae Montesinos, Cleef & Sýkora 2021, del sur (sector de Arequipa). La alianza pertenece al orden Saxifragetalia magellanicae Galán & Cáceres in Galán, Rosa & Cáceres 2002, de la clase de vegetación rupícola de pequeñas fisuras ricas en pteridófitos Woodsio montevidensis-Cheilanthetea pruinatae Galán, Campos, Linares, Montoya, Torres & Vicente 2021.
Conclusión
Como conclusión a este estudio, damos a conocer el esquema sintaxonómico de las unidades de vegetación tratadas.
CLEMATIDO PERUVIANAE- BACCHARITETEA LATIFOLIAE Galán,
Sánchez, Montoya, Linares, Campos & Vicente 2015
+ Mutisio acuminatae-Baccharitetalia latifoliae Galán & Cáceres in Galán, Rosa & Cáceres 2002
* Clinopodion bolivianae Seibert 1993 nom. mut. nov. [nombre original: Saturejion bolivianae (Seibert 1993: 278)]
1. Berberido bolivianae-Flourensietum polycephalae Galán, Linares, Montoya & Vicente ass. nov.
* Kageneckio lanceolatae-Tecomion sambucifoliae Galán, Linares, Montoya & Vicente all. nov.
2. Schino mollis-Prosopidetum andicolae Galán, Linares, Montoya & Vicente ass. nov.
II. ACACIO MACRACANTHAE- PROSOPIDETEA PALLIDAE Galán 1999
+ Acacio macracanthae-Prosopidetalia pallidae Galán 1999
* Tecomion fulvae Galán & Cáceres in Galán 1999
3. Schino mollis-Prosopidetum calderensis
Galán, Linares, Montoya & Vicente ass. nov.
HYMENOXYO ROBUSTAE- DISTICHLIETEA SPICATAE Galán,
Linares, Montoya & Vicente cl. nov.
+ Hymenoxyo robustae-Distichlietalia spicatae Galán, Linares, Montoya & Vicente ord. nov.
* Salicornio cuscoensis-Distichlion spicatae
Galán, Linares, Montoya & Vicente all. nov.
4. Salicornietum cuscoensis Gutte & Müller 1985
5. Hymenoxyo robustae-Distichlietum spicatae Galán, Linares, Montoya & Vicente nom. nov. [Distichlietum spicatae Gutte & Müller 1985 nom. corr. [nombre original corregido: Distichlietum humilis (Gutte & Müller, 1985: 405), nom. inept.], non Distichlietum spicatae (López Guillén & Rivas Goday 1952: 606]
IV. XYRIDO CAROLINIANAE-TYPHETEA DOMINGENSIS O. Bolòs, Cervi & Hatschbach 1991
+ Schoenoplectetalia olneyi-americani Galán, Linares, Campos & Vicente 2009
* Schoenoplection tatorae Galán, Campos, Linares, Montoya, Torres & Vicente 2021
6. Schoenoplectetum tatorae Galán 1995, nom. mut. Galán, Campos, Linares, Montoya, Torres & Vicente 2021
7. Schoenoplectetum badii Galán, Linares, Montoya & Vicente ass. nov.
V. CALAMAGROSTIETEA VICUNARUM
Rivas-Martínez & Tovar 1982
+ Coreopsietalia fasciculatae Galán, Campos, Linares, Montoya, Torres & Vicente 2021
* Plantaginion lanuginosae Gutte 1986 nom. mut. Galán, Campos, Linares, Montoya, Torres & Vicente 2021
8. Poo candamoanae-Festucetum humilioris Galán, Linares, Montoya & Vicente ass. nova
WOODSIO MONTEVIDENSIS- CHEILANTHETEA PRUINATAE Galán,
Campos, Linares, Montoya, Torres & Vicente 2021
+ Saxifragetalia magellanicae Galán & Cáceres in Galán, Rosa & Cáceres 2002
* Saxifragion magellanicae Galán & Cáceres in Galán, Rosa & Cáceres 2002
Woodsio montevidensis-Begonietum veitchii
Galán, Linares, Montoya & Vicente ass. Nova

