











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCTION
Maize (Zea mays L.) presents an extraordinary genetic and morphological diversity, product of a multiple and independent domestication (Márquez-Licona et al., 2021). Maize along with rice and soybeans, are strategic to meet the growing demand for food (Melendez, et al., 2021). Also is an important crop in the Ecuadorian Highlands because it is an indispensable food in the diet of the residents and is one of the main sources of economic income for the small and medium producer (Aguirre et al., 2022). The planted area is 330,337 hectares with a production of 1324147 t (INEC, 2018; FAO, 2018). The most widely cultivated types of maize in Ecuador are distributed according to the customs and preferences of farmers, with hard corn being the predominant in the coast region and soft corn in the highland’s region (Yánez et al., 2013; Bravo & León, 2013).
The purple corn plant has a dark-colored cob; due to this, in some places it is known as black corn; this color occurs because the rachis and pericarp that cover the grain have a high content of anthocyanins, pigment that is in high demand in the food, cosmetic and pharmaceutical industries (Lao et al., 2017; Somavat et al., 2018).
On the other hand, the low yield of black corn at the national level makes it necessary to generate varieties with higher yield; thus, the purpose of the genetic improvement programs is to identify superior lines for the formation of new hybrids or synthetics (Salinas et al., 2016). To obtain corn hybrids, it is necessary to obtain controlled self-pollinated lines. A line is produced by self-fertilization and selection until apparently homozygous plants are obtained after five to seven generations, causing reduction in the vigor of the plant (Huerta-Santoyo et al., 2019).
A self-fertilized line is the descendant population obtained from the self-pollination of a plant in each generation. Self-fertilization is the union of male and female gametes from the same plant with the aim of forming a new individual with increased proportion of alleles in homozygous state in relation to the previous generation (Narro & Arcos, 2015); however, corn germplasm in the Ecuadorian Sierra has low tolerance to inbreeding, especially in starchy or mild cultivars. In this regard, to obtain commercially competitive hybrids it is necessary to identify outstanding progenitor lines in early stages (Noor et al., 2013; Trachsel et al., 2016).
The early test consists of identifying and selecting outstanding lines to be used for genetic improvement and forming possible combinations to take advantage of heterosis in some crosses (Huerta-Santoyo et al. 2019). On the other hand, when there are genes that determine desirable quantitative characteristics, an early test allows to concentrate them by gradual selection and recombination to generate superior populations with high yield potential (Rendón-Aguilar et al., 2015). Regarding the use of the early generation test of inbred lines, some researchers consider that these can be selected for their performance, since it is positively correlated with their general combining ability (GCA), with values larger than 0.5 (Salinas et al., 2016). Genetic depression can results in low correlations with performance at advanced levels of inbreeding, but the initial ACG of the lines is inheritable (Noor et al., 2013).
The objective of this research was to evaluate the yield potential of S1 lines of black corn through early testing, through the identification of lines with greater productive potential and the selection of superior genotypes of black corn to produce advanced generations.
MATERIALS AND METHODS
Plant material and experimental design
The experiment was conducted at “Tunshi” Experiment Station (01°45′S; 78°37′W), with an annual temperature of 14 °C; annual precipitation between 450 and 500 mm and relative humidity of 60%; it is located at the Licto parish, Chimborazo province, Ecuador. A complete random block design with four replications was used. The experimental unit consisted of a 4.4 m2 (5.5 × 0.80 m) plot, where two seeds were placed per site, 0.50 m separated. Twenty-four treatments were evaluated, each treatment consisted of an S1 line of black corn (Table 1).
Agronomic management
The soil was prepared using one pass with a moldboard plow, two passes with a disc harrow plow, and two passes with a vibro-cultivator. Manual seeding was done with a rate of 35 kg/ha of seed. After soil analysis, fertilization was uniformly applied as 80 kg N/ha and 60 kg P2O5/ha, 50% of the nitrogen was applied before planting and the remaining 50% at hilling, 45 days after planting. Additionally, a skimming was performed 20 days after planting and two hilling 45 and 90 days after planting.
Furrow irrigation was applied every 15 days; in addition, rainfall of 514 mm was present during the crop cycle that favored the development of corn in the final stages. Weeds were controlled preemergently in humid soil with Atrazine herbicide (2 kg/ha), also, mechanical/manual weed removal was carried out in every hilling. Agrotis ipsilon (Hufnagel) (Lepidoptera: Noctuidae) was controlled with Lambdacialotrina insecticide (20 cm3/ha), applied in the base of plants base.
Table 1 Genealogy and origin of each black maize line evaluated through the early test. Tunshi, Ecuador
| TREATMENTS* | ORIGIN OF THE COLLECTIONS | |
| 1 | CDE-024 (1) | Gualabí alto locality, El Lago parish, Otavalo canton, Imbabura province. Cob 1. |
| 2 | CDE-024 (2) | Gualabí alto locality, El Lago parish, Otavalo canton, Imbabura province. Cob 2. |
| 3 | CDE-024 (3) | Gualabí alto locality, El Lago parish, Otavalo canton, Imbabura province. Cob 3. |
| 4 | CDE-025 (1) | Pibaringí locality, Los Lagos parish, Otavalo canton, Imbabura province. Cob 1. |
| 5 | CDE-025 (2) | Pibaringí locality, Los Lagos parish, Otavalo canton, Imbabura province. Cob 2. |
| 6 | CDE-025 (3) | Pibaringí locality, Los Lagos parish, Otavalo canton, Imbabura province. Cob 3. |
| 7 | CDE-025 (4) | Pibaringí locality, Los Lagos parish, Otavalo canton, Imbabura province. Cob 4. |
| 8 | CDE-035 (1) | Tandalibí locality, Alaquez parish, Latacunga canton, Cotopaxi province. Cob 1. |
| 9 | CDE-042 (1) | Guapulo locality, Pujilí parish, Latacunga canton, Cotopaxi province. Cob 1. |
| 10 | CDE-042 (2) | Guapulo locality, Pujilí parish, Latacunga canton, Cotopaxi province. Cob 2. |
| 11 | CDE-056 (1) | Pilgacho Grande locality, Mashingui parish, Pedro Moncayo canton, Pichincha province. Cob 1. |
| 12 | CDE-056 (2) | Pilgacho Grande locality, Mashingui parish, Pedro Moncayo canton, Pichincha province. Cob 2. |
| 13 | CDE-056 (3) | Pilgacho Grande locality, Mashingui parish, Pedro Moncayo canton, Pichincha province. Cob 2. |
| 14 | CDA-058 (1) | Loma Llushapa locality, Ushapa parish, Saraguro canton, Loja province. Cob 1. |
| 15 | CDA-058 (2) | Loma Llushapa locality, Ushapa parish, Saraguro canton, Loja province. Cob 2. |
| 16 | CDA-058 (3) | Loma Llushapa locality, Ushapa parish, Saraguro canton, Loja province. Cob 3. |
| 17 | CDA-059 (1) | Centro Saraguro locality, Saraguro parish, Saraguro canton, Loja province. Cob 1. |
| 18 | CDA-059 (2) | Centro Saraguro locality, Saraguro parish, Saraguro canton, Loja province. Cob 2. |
| 19 | CDA-059 (3) | Centro Saraguro locality, Saraguro parish, Saraguro canton, Loja province. Cob 3. |
| 20 | CDA-059 (4) | Centro Saraguro locality, Saraguro parish, Saraguro canton, Loja province. Cob 4. |
| 21 | CDA-059 (5) | Centro Saraguro locality, Saraguro parish, Saraguro canton, Loja province. Cob 5. |
| 22 | CDA-059 (6) | Centro Saraguro locality, Saraguro parish, Saraguro canton, Loja province. Cob 6. |
| 23 | CD-072 (1) | Loma Cruz locality, Matriz parish, Chillanez canton, Bolívar province. Cob 1. |
| 24 | CD-072 (2) | Loma Cruz locality, Matriz parish, Chillanez cantonb, Bolívar province. Cob 2. |
*The first six characters belong to the code assigned in the original cycle, the number in parentheses indicates the cob number from which the seed was extracted.
Agronomic variables
The following variables were evaluated: Days to Male Flowering (DMF), counting the number of days from planting until 50% of plants in the experimental unit released pollen; Days to Female Flowering (DFF), registering the number of days from planting until 50% of plants in the experimental unit exposed stigmas at least 2 cm long; Floral Asynchrony (FA), corresponds to the interval between days to male and female flowering, these data are not presented in table 2 as they were calculated based on DMF and DFF; however, it is mentioned and explained in the results section; Plant Height (PH) (cm), measured from the plant base to the branching point of tassel, this value was registered after flowering in 10 plants randomly; Cob Height (CH) (cm), measured from the plant base to the insertion node of the upper ear, this value was registered after flowering in 10 plants randomly; prevalent Leaf Disease (LD), assessed 45 days after female flowering; Tassel Length (TL) (cm), the distance between the first and the last primary branch of 10 plants randomly, after milky state (Centro Internacional de Mejoramiento de Maíz y Trigo [CIMMYT], 2017; Salinas et al., 2016); Cob Rot percentage (CR), qualifying all the cobs harvested in each experimental unit, according to the scale used by Briones-Reyes et al. (2015) with modifications, where: 1 = 0-20; 2 = 21-40; 3 = 41-60; 4 = 61-80; 5 = 81-100% of grains with symptoms and the average was obtained in each plot based with the equation:
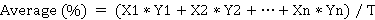 (1)
(1)
Where: X is the number of cobs at each value on the scale, Y is the corresponding average value and T is the total number of cobs.
Other traits evaluated were the Number of Grains per Row (NGR), averaging the number of grains of three rows in 10 cobs taken randomly from each plot; Row Number per Cob (RNC), as the average number of rows of 10 ears; Cob Length (CL) (cm) from the insertion of the peduncle to its apex, averaging 10 cobs; Cob Diameter (CD) (cm) measured with a caliper at the central part of 10 upper cobs taken randomly; De-grained Cob Diameter (DCD) (cm) measured with a caliper at the central part of 10 de-grained cobs taken randomly; 100- Grain Weight (GW) (g), determined after harvest and threshing 5 cobs and recording the weight of 100 grains taken randomly; Grain Yield (GY) from the whole plot and adjusting to 14% humidity, then it was weighed on a digital scale and the result expressed as t/ha (Salinas et al., 2016).
The following formulas were applied to calculate the inbreeding of each variable (Escorcia-Gutierrez et al., 2010):
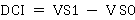 (2)
(2)
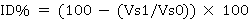 (3)
(3)
where DCI corresponds to the difference caused by inbreeding, ID is the inbreeding depression in percentage, VS1 is the value of the self-fertilized population and VS0 is the value of the original population.
Statistical analysis
Data were subjected to analysis of variance at P ≤ 0.05 after verifying the assumptions of normal distribution and homogeneity of variances. When these assumptions were not fulfilled, transfor-mations were performed with the ln (x+1) function. Means were compared by the Tukey test at the 95% confidence level. The statistical analysis was performed with the INFOSTAT statistical program (Di Rienzo et al., 2017).
RESULTS AND DISCUSSION
The comparison tests of means of variables that presented significant differences (P ≤ 0.05) are presented in Table 2. The lines presented differences (P ≤ 0.05) in the variables Days to Male Flowering (DMF), Days to Female Flowering (DFF), Plant Height (PH), Cob Height (CH), Leaf Disease (LD), Cob Length (CL), weight of 100 Grains (GW) and Grain Yield (GY); while variables Panicle Length (PL), Cob Rot (CR), Number of Grains per Row (NGR), Number of Rows per Cob (NRC), Cob Diameter (CD); and, De-grained Cob Diameter (DDC) were statistically equal (P > 0.05) between lines.
It was observed that line 24 presented the highest DMF (135.7 days), while lines 17 and 18 presented the lowest values (108.0 and 107.3 days, respectively). Inbreeding depression obtained an average gain of 2.12%, showing a slight delay of anthesis during this cycle.
Line 24 presented the highest DFF value (142.3 days), while line 13 showed the lowest value (114 days). The percentage of inbreeding depression shows that this variable had an average reduction of 2.54.
The mean DMF of the original accessions (S0) used to calculate the percentage of inbreeding was 115.69 days, DFF was 129.81 and floral asynchrony (FA) was 14.13. The mean data of the self-fertilized accessions (S1) for DFM and DFF were 119.64 and 127.60 days, respectively, and the FA was 7.96 days, evidencing a reduction of 56.33%.
Early testing on inbred maize lines enables to advance the selection process faster (Bekavac et al., 2008). The differences in DMF and DFF obtained in our study are due to the genetic variability of the original populations, which precede populations collected in different locations in the Ecuadorian highlands. Our results are similar to those obtained by Cervantes-Ortiz et al. (2016), who indicated that the differences in days to flowering are due to the genetic diversity of the materials studied. On the other hand, in the corn populations studied by Bekavac et al. (2008), it was determined that this characteristic presented highest genetic varia-bility; while Salinas et al. (2016) pointed out that the variability in flowering is due to the allogamous nature of corn to ensure pollination; they also showed that the differences between lines for the variables days to male and female flowering are due to genetic diversity of lines, concluding that the use of the early test in S1 lines is appropriate to identify and select superior genotypes. Kliebenstein (2010) pointed out that differences in flowering time in inbred lines are the result of cumulative effects of many loci.
For their part, Tadeo-Robledo et al. (2016), when evaluating productivity in corn hybrids, they found significant differences in male flowering days.
In PH variable, line 16 presented the highest average (237.7 cm), while line 9 presented the lowest average for this variable (146.3 cm). In the CH variable, it was observed that lines 23 and 16 presented the highest average (148.2 and 143.8 cm respectively), while line 9 presented the lowest value (69.77 cm). The percentage of inbreeding depression showed a reduction of 32.88%. The data obtained in CH and PH variables are similar to the data obtained by Salinas et al. (2016), who showed that the evaluated genotypes presented a lot of variability, both in plant height and cob height, so that lines with better plant and cob height can be selected. Some studies (Arellano et al., 2003; De la Cruz-Lazaro et al., 2009; Tadeo-Robledo et al., 2016; Vega et al., 1998) evaluated corn genotypes, identifying in a short time, lines with good plant height and cob height.
Table 2 Comparison of means using Tukey test 5% in the agronomic variables evaluated by the early testing on S1 lines of black corn. Tunshi, Ecuador
| Treatments | Variables | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| DMF | DFF | PH | CH | LD | CL | GW | GY | ||
| 1 | CDE-024 (1) | 133.70 ab | 140.00 ab | 210.90 abc | 127.40 abcd | 2.00 cd | 10.94 ab | 74.23 a | 1.01 c |
| 2 | CDE-024 (2) | 132.30 ab | 139.00 ab | 230.50 abc | 128.60 abcd | 1.67 d | 11.78 ab | 53.51 ab | 1.34 c |
| 3 | CDE-024 (3) | 127.00 abc | 133.30 abc | 227.50 abc | 130.10 abc | 2.33 bcd | 11.61 ab | 58.25 ab | 1.08 c |
| 4 | CDE-025 (1) | 126.30 abcd | 130.70 abcdef | 199.20 abc | 121.20 abcd | 2.67 abcd | 10.50 ab | 48.09 ab | 1.21 c |
| 5 | CDE-025 (2) | 125.30 abcde | 130.30 abcdef | 228.00 abc | 131.60 abc | 2.67 abcd | 10.90 ab | 48.15 ab | 1.14 c |
| 6 | CDE-025 (3) | 122.00 abcdef | 130.30 abcdef | 218.70 abc | 126.90 abcd | 2.33 bcd | 12.60 ab | 50.12 ab | 1.04 c |
| 7 | CDE-025 (4) | 125.30 abcde | 132.70 abcd | 176.80 abc | 101.80 abcd | 2.00 cd | 10.33 ab | 31.92 b | 0.84 d |
| 8 | CDE-035 (1) | 120.30 abcdef | 130.30 abcdef | 149.80 bc | 81.100 bcd | 2.33 bcd | 11.00 ab | 55.59 ab | 0.31 d |
| 9 | CDE-042 (1) | 124.70 abcde | 130.30 abcdef | 146.30 c | 69.77 d | 2.67 abcd | 8.29 b | 40.82 ab | 0.75 d |
| 10 | CDE-042 (2) | 118.70 bcdef | 128.30 bcdef | 153.50 abc | 79.27 cd | 2.67 abcd | 9.33 b | 45.72 ab | 0.76 d |
| 11 | CDE-056 (1) | 110.70 def | 118.70 fg | 180.10 abc | 94.10 abcd | 3.00 abc | 9.00 b | 58.40 ab | 1.00 c |
| 12 | CDE-056 (2) | 112.30 cdef | 119.70 efg | 174.20 abc | 103.10 abcd | 2.00 cd | 10.22 ab | 53.89 ab | 1.01 c |
| 13 | CDE-056 (3) | 109.30 ef | 114.00 g | 198.40 abc | 100.40 abcd | 3.33 ab | 11.78 ab | 59.40 ab | 1.57 bc |
| 14 | CDA-058 (1) | 127.00 abc | 131.30 abcde | 212.80 abc | 133.30 abc | 3.00 abc | 10.67 ab | 48.86 ab | 1.14 c |
| 15 | CDA-058 (2) | 123.30 abcdef | 129.00 bcdef | 232.20 abc | 140.30 ab | 3.67 a | 11.50 ab | 55.64 ab | 2.00 a |
| 16 | CDA-058 (3) | 120.30 abcdef | 129.30 bcdef | 237.70 a | 143.80 a | 3.33 ab | 10.34 ab | 48.32 ab | 2.08 a |
| 17 | CDA-059 (1) | 108.00 f | 121.00 cdefg | 222.60 abc | 121.80 abcd | 3.33 ab | 10.67 ab | 56.96 ab | 1.15 c |
| 18 | CDA-059 (2) | 107.30 f | 121.00 cdefg | 203.40 abc | 112.30 abcd | 3.33 ab | 10.94 ab | 47.27 ab | 1.73 b |
| 19 | CDA-059 (3) | 110.70 def | 122.30 cdefg | 212.20 abc | 129.30 abcd | 2.00 cd | 10.15 ab | 58.71 ab | 1.59 bc |
| 20 | CDA-059 (4) | 111.00 cdef | 120.30 defg | 225.40 abc | 124.00 abcd | 3.00 abc | 9.33 b | 53.01 ab | 1.71 b |
| 21 | CDA-059 (5) | 110.30 def | 119.30 efg | 207.70 abc | 114.70 abcd | 2.67 abcd | 10.39 ab | 52.54 ab | 1.52 bc |
| 22 | CDA-059 (6) | 111.30 cdef | 124.00 cdefg | 219.70 abc | 131.70 abc | 2.33 bcd | 10.89 ab | 53.38 ab | 1.95 a |
| 23 | CD-072 (1) | 133.00 ab | 139.70 ab | 221.60 abc | 148.20 a | 2.33 bcd | 13.83 a | 61.61 ab | 0.91 d |
Different lowercase letters indicate significant differences between treatments (Tukey P ≤ 0.05).
For PL variable, it was observed that line 15 presented the highest average (18.53 cm), while line 9 showed the lowest value (10.20 cm). The average value of the percentage of inbreeding depression presented a reduction of 2.67%.
The prevalent leaf disease (LD) was Helmin-thosporium turcicum; when evaluating this variable, it was observed that line 15 showed the highest value (3.67); on the contrary, line 2 presented the lowest value for this variable (1.67). The average value of the percentage of inbreeding depression was 42.75%. In our study, the average value of the percentage of inbreeding depression for LD caused by Helminthosporium turcicum, was the highest of all the agronomic variables evaluated. For their part, Soto-Rocha et al. (2019) when evaluating the impact of foliar diseases on the yield of corn genotypes, they found that H. turcicum affected the hybrids studied, so 56% obtained yields less than 5.5 t/ha. Barrón (2008) and Ramirez-Díaz et al. (2019) pointed out that in corn cultivation the severity of foliar diseases has increased due to the accelerated growth of the cultivated area and the use of susceptible genetic material.
In the variable CR caused by Fusarium spp., it was observed that the lines presented inbreeding depression (38.41%). For the RNC variable, it was observed that the average value of the percentage of inbreeding depression indicated a reduction of 3.69%. In the NGR variable, it was observed that the average value of the percentage of inbreeding depression indicated a reduction of 9.98%, also evidencing a slight depression. The CR variable had a high inbreeding depression value. Incidence of molds and mycotoxins caused by Fusarium spp., affect the corn grain, decreasing production and altering its nutritional value, because they cause the death of the embryo in the grain with the consequent economic losses that this entails (Chavarri et al., 2017).
In the CL variable it was observed that line 23 presented the highest value (13.83 cm), while the lowest values were presented by lines 9, 10, 11 and 20, with averages of 8.29; 9.33; 9.00 and 9.33 cm, respectively. Regarding inbreeding depression, a 14.07% reduction was also observed, indicating that there was a decrease in relation to the average of the previous cycle. The results obtained in CL variable, are similar to those obtained by Salinas et al. (2016), when evaluating 80 S1 lines of corn, who found wide variability, because only 15 lines presented cob length similar to the control. Mendes et al. (2008), found that cob length variable was greater than in the original population, when they carried out recurrent selection work on S2 progenies of corn.
For the CD variable, the percentage of inbreeding depression indicated a reduction of 11.05%. This meant that there was a decrease in the self-fertilized line in relation to the original line. The DCD variable showed 21.74% of inbreeding depression, which indicates a considerable decrease in this variable. In our study, no significant differences were found for CD, these results agree with those obtained by Salinas et al. (2016) in the same variable. Similarly, no significant differences were obtained for the NGR, results similar to those found by Salinas et al. (2016). However, Mendes et al. (2008) found that the number of grains per row was greater than that of the original population in S2 progenies. In the variable RNC, in our study there were no significant differences; however, Salinas et al. (2016) found differences, with values ranging from 28.3 to 40 grains per row.
For the GW variable, it was observed that line 1 presented the highest average (74.23 g), while lines 7 and 24 presented the lowest values with averages of 31.92 and 31.05 g, respectively. The percentage of inbreeding depression showed a reduction of 23.67%.
In the GY variable, it was observed that the percentage of inbreeding depression showed a reduction of 41.26%. In addition, lines 15, 16 and 22 presented the highest returns with 2.00; 2.08 and 1.95 t/ha respectively. In our study, the GY variable presented significant differences between lines, similar to that reported by Salinas et al. (2016). In this first generation of self-fertilization, high heterogeneity was observed, which is reduced with the advance in the inbreeding process and the reduction in the degree of heterozygosity. In this regard, Mendes et al. (2008) and Mengesha (2013) say the best grain yield can be obtained in the first selection cycles, but then it decreases, as the inbreeding process progresses. As the degree of inbreeding increases, the lines become more homogeneous but the variability between them increases. The variability observed in the variables is an important factor for the selection process of superior lines, especially those that are directly related to yield (Tucker et al., 2020). Esquivel et al. (2009) and Edmeades et al. (2017) indicated that, by applying early tests on maize germplasm, it is possible to identify outstanding genotypes and also have genes for desirable quantitative traits, which later through progressive recombination and selection can obtain higher populations.
CONCLUSIONS
The use of early test allows to identify S1 lines of black corn with outstanding yields, which would allow in the future to select lines with less depressive effects on the yield produced by inbreeding. All the evaluated variables presented inbreeding depression, with the exception of days after male flowering, which presented a slight increase. The floral asynchrony suffered a reduction in relation to the asynchrony of the original cycle, since this is a measure of adaptive value and the accessions are still in the process of adapting to their new environment. The variable that presented the highest percentage of inbreeding depression was prevalent leaf disease.
Early testing is a technique to exclude lines with undesirable agronomic characteristics and low yield potential.
It is recommended that the lines selected in this study and that have less depressive effects on yield be used to continue with the improvement processes of black maize in Ecuador.

