











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCTION
The Citrus genus belongs to the Aurantioideae subfamily of Rutaceae (Gonzales & Tullo, 2019; Reuther & Webber, 1967). Citric species have several origins, for example sweet oranges and tangerines are from China, and lemon are from India (Talon et al., 2020). However, there is new evidence indicating that other citrus species are native to Australia and surrounding areas (Beattie et al., 2008). Although citric fruits can be grown worldwide, they are usually cold-sensitive, so ideal growing conditions are found in tropical and subtropical regions along the equatorial line (Talon et al., 2020).
Citric fruits represent the main fruit crop with the largest surface and production area (Orrego et al., 2020), being the species of the greatest economic importance: Clementin (Citrus x clementina), lemon (C. x lemon), grapefruit (C. x paradisi), lime (C. aurantifolia, C. x latifolia) sweet orange. (C. x sinensis) and tangerine (C. x reticulata) (Wu et al., 2018). Out of this group, orange contributes to more than half of the production in citric fruits, with 130 million metric tons produced during the period 2018-2019 (World Citrus Organization, 2019).
Brazil is known as the first producer country of (C. x sinensis), with a production of 17’073.593 t.ha-1 and a yield of 28.96 t.ha-1 (FAOSTAT, 2019). Meanwhile in Ecuador, the production of orange and lemon in fresh fruit was 133.688 t, and 19.672 t, respectively. Manabi province is the main producer of lemon with 5.510 t.ha-1, on a planted area of 1.348 ha as monoculture, according to data reported by National Institute of Statistics and Census (INEC, 2019).
According to Zayas (2019), the main element that allows a feasible development of citric production is the predominant climate, for that reason the planting of citric fruits depends on the season of the year that promotes its vegetation, being the tropical and subtropical climate the most suitable, due to its seasonality. One of the setbacks faced by fruit trees is usually associated with the production chain, given by the variability of supply and demand in a season (González, 2009).
Castelló (2019), when referring to the law of supply and demand, says that it is unappealable, since there is an excessive supply of a product, the price to the consumer is reduced significantly, affecting the producers' economy. In addition, Westreicher (2019) mentions that the characteristics of excess supply emphasizes that there is an imbalance in the market, due to the existence of goods in abundance, causing price reductions, thus affecting the producer's economy.
Regarding Ecuador, the province of Manabí, in the 2018 registered that the months with the lowest production of subtle lemon were from July to November (Valarezo et al., 2020), reason for which prices were higher, i.e., the cost of the 35 kg bag was approximately USD 35. On the other hand, MAG (2018) mentions that from August to November the price of lemon ranged from USD 17 to USD 32.
Because of the latter, certain citrus producers detailed the price variations of oranges and lemons in the province of Manabi in the first period of 2021; thus, Bazurto (2021) mentioned that orange is scarce and expensive during November 2020 to February 2021 and its prices ranged from USD 8 to 12, while the price of the 35-kg bag of lemon was USD 4 to USD 5. As of July 2021, prices for tangerine and orange were USD 3 and 5, respectively. However, lemon was scarce and the price of the 35-kg bag increased to USD 40 and lemon to USD 18. The price of the 35-kg lemon bag from January to March 2021 was USD 5 to 8 and from April to May USD 10, while in June it increased to USD 20. From July to September, it has remained at USD 35 to 30. Tuárez (2021) is a producer who mentions that on his farm he stopped harvesting since June 2021, due to the low flowering.
PBZ contributes to floral uniformity and production (Michelini & Chinnerry, 1988; Michelini & Chinnerry, 1989). This element can inhibit the synthesis of gibberellins, favoring the expression of the FLOWERING LOCUS T (FT) gene in charge of floral induction (Nakagawa et al., 2012), which generates the expression of flowering genes (Corbesier et al., 2007; Zeevaart, 2008; Muñoz et al., 2012). PBZ has also been used as an effective protector, because it helps to overcome environmental stress in plants (Javid et al., 2011) and it has been related to the increase in the content of antioxidant enzymes by providing protection to the plant; another use is to promote a greater production of the crops (Özmen et al., 2003). Finally, some research has related it to dissipating the stress caused by salinity in barley and wheat (Javid et al., 2011). For this reason, the objective of the research was to develop an exhaustive search for information regarding the importance of PBZ in the out-of-season production of citric fruits.
FLOWERING OF CITRIC FRUITS
It is important to note that in fruit trees such as citrus fruits, a phenomenon called productive alternation can occur, i.e. years “on” (high production) and years “off” (low production) (Angel, 2021), related to the inhibition of flowering as a result of genetic, nutritional, and hormonal factors (Lavee, 2007), photoperiod, vernalization and environmental temperatures, which suppress the expression of the gene in charge of flowering (FT) (Andrés & Coupland, 2012), as a result of unfavorable conditions.
The floral behavior of fruit trees differs from other species, because their flowering may be induced by environmental signals; however, it may be controlled locally as a result of the presence of fruits in development from the previous cycle (Martínez-Fuentes et al., 2010), Consequently, it is possible to implement agricultural strategies to optimize resources and to have a better production distribution over the time (Martínez-Alcantara et al., 2015)
The limitation of the number of flowers results from the large quantity of fruits harvested, called productive alternation. These effects may be related to poor nutrition, causing a decrease in reserves before floral differentiation, or the floral inhibition caused by gibberellin generated in fruits (Zeng et al., 2019).
To prevent an excessive intensity of flowering that affects the curd of fruits and significantly reduces the harvest, gibberellic acid is applied as a hormonal treatment (floral inhibition agent) to promote good flowering and an adequate curd of fruits (Nishikawa et al., 2017).
PBZ treatments can only stimulate flowering in trees with a limited number of fruits (years off), i.e., those expressing CiFT2 genes responsible for floral induction. However, in those trees with a high production (years on), hormonal methods will have no results (Martínez-Fuentes et al., 2013).
METHODS OF PRODUCTION REGULATIONS IN CITRUS
The main treatments of flowering regulation of citrus fruits are: water stress, threshing, and fruit pruning.
Water stress. According to Delgado (2018) when referring to subtle lemon, when it was subjected to an irrigation suspension from 30 to 45 days, the number of buds and inflorescence increases; however, enhancing its effect can be associated with other strategies such as the removal of flowers and fruits to reduce competition.
Threshing. It consists of removing the bark of the trunk or branch in a circular way, and it involves tissues such as phloem, favoring the accumulation of carbohydrates and the reduction of growth (Cervantes et al., 2018).
Fruit pruning. It is one of the main activities in citrus fruits that favors the quantity, quality and size of fruits (Artavia-Mejia, 2019), due to an optimal distribution and balance between vegetation and fruit. The photosynthetic process is improved, allowing continuous production (Angel, 2021).
Hormones were discovered in the early 20th century, and have been used as an agronomic instrument, mainly in fruticulture (Lal et al., 2017). Phytohormones act at the cellular level, and occur at low concentrations (Kumar et al., 2021), and currently 5 groups of phytohormones are known (Aremu et al., 2020). These include: Abscisic acid, gibberellic acid, indoleacetic acid, cytokines and ethylene (Kwon et al., 2020). Because of phytohormones, synthesis products (industrial) appeared, and the term ‘growth regulator’ was coined (Lal et al., 2017), these are much more effective than their natural analogues; for this reason, it is essential to know aspects such as: ration, susceptibility of variety, plant status, etc. (Kwon et al., 2020).
Plant regulators are used in growth and development processes, through modifications of associated biochemical and physiological techniques, of which PBZ is considered one of the most variable plant growth retardants, able to limit vegetative growth to stimulate flowering in several fruit trees (Kumar et al., 2021) including citrus (Aron et al., 1985). One of the options suggested by Rani et al. (2018) is to perform pruning and apply PBZ; this process is used in many fruit crops to control tree vigor and improve productive efficiency, while Hegde et al. (2018) mentions its effectiveness in reducing plant height, because PBZ restricts the synthesis of gibberellins in subapical meristem, limiting elongation and cell division, resulting in decreased growth of shoots.
IMPORTANCE OF TRIAZOLES IN CITRUS PRODUCTION
Rugeles (2017) emphasizes that triazoles have become the most used universal group for grain protection, encompassing approximately 60% of the world market. One of the benefits it brings is its wide spectrum effectiveness of fungal diseases, such as Oidium sp., Puccinia sp., Phytophthora infestans, etc.; this compound actions against Deuteromycetes - Ascomycetes and Basidiomycetes groups, because they can stop the sterol synthesis (Chaney, 2005).
Triazoles have a systemic action. They move by means of the xylem (acropetal action) i.e., from the base toward the apex and by leaves (translaminar circulation) (López, 2019). As a result of their systemic and acropetal movement, triazoles preserve growth sites (Rugels, 2017). This substance has certain useful attributes linked to the adjustment of plant growth (Presser, 2016). It improves the chlorophyll content in the tissues, increases the assimilation phase and photosynthesis, improving the filling period of fruits and grains (López, 2019).
According to Davis et al. (1988), the main use given to PBZ is to control or reduce the stems height growth and outbreaks (Phadung et al., 2011), in citrus patterns (Hadlow, 1988) due to the decrease in the biosynthesis of gibberellin (Hartmann et al., 2011), resulting in a reduction in vegetative growth (Chaney, 2005), increasing the total carbohydrate content which results in flowering (Yeshitela et al., 2004).
PBZ has also been used to increase the number of flowers in citrus crops as stated by Munoz et al. (2012) who applied 40 mg L-1 of gibberellic acid and 2000 mg L-1 of PBZ in orange plants, which were sprayed using 10 L per tree, resulting in a 72% reduction in the number of flowers per 100 knots when applying gibberellic acid, and increasing the growth of vegetative buds. However, PBZ treatments managed to increase the number of flowers per 100 knots by 123% and the number of germinated buds by 74%, and the number of vegetative shoots was reduced.
Regarding the latter, it can be mentioned that triazoles have the capacity to alter the hormonal balance of plants, being the most important gibberellin, abscisic acid and cytokines (Hajihashemi et al., 2007). This substance can promote the increase in the synthesis of abscisic acid, causing morphological modifications of the leaf, such as the reduction in the number of stomas and can increase the synthesis of cytokines, favoring root growth, providing greater protection against environmental stress (Chaney, 2005)
EFFECT OF THE USE OF PHYTOHORMONES AND PBZ ON FORCED CITRUS PRODUCTION
Triazole compounds such as PBZ inhibit the synthesis of gibberellins and auxins (AIA), shortening cell elongation and cell division (Hartmann et al., 2011). PBZ has certain properties as a growth retardant (Lewis et al., 2016), because it specifically inhibits the three steps in the oxidation of the precursor (Hedden & Graebe, 1985) of the gibberellic acid from ent-kaurene to ent-kaurenoic acid (Hedden & Graebe, 1985; Rademacher, 2016) by disabling cytochrome P-450-dependent oxygenase (Hedden & Graebe, 1985). However, the effect of PBZ can be modified through exogenous application of gibberellic acid (Lever, 1986).
Meanwhile, abscisic acid is synthesized by isoprenoid like gibberellins, however, these two compounds generally have opposite physiological behaviors. Abscisic acid is a phytohormone related to the inhibition of several processes, improving acclimatization and environmental stress in plants (Mackay et al., 1990; Assmann, 2010). The effect of PBZ on abscisic acid is that it acts as a stress protection (Fletcher & Hofstra, 1988), because this helps to improve the concentrations of abscisic acid in leaves, influencing the closure and opening of the stomata, reducing the water impairments by means of transpiration, it also decreases the growth of outbreaks, reduces the surface area for breathability and increases the root index for greater water absorption (Desta & Amare, 2021).
Rook et al. (2001) emphasizes the importance of abscisic acid in the process of regulating the metabolic pathways of sugars, increasing stimulation in tissue response, which may be related to what was proposed by Flórez & Pereira (2008) who mentioned that the opposite levels of abscisic acid and indoleacetic acid accelerate the floral development in Solidago x luteus. According to Bhattacharya (2019), cytokines participate in many aspects of growth and development, Jordán & Casaretto (2006) mention that they promote cell division, stimulate the growth of outbreaks, and lateral and dormancy buds activate them, and promote the development of chloroplasts.
Triazoles stimulate the synthesis of cytokines by improving the development of chloroplasts, avoiding the degradation of chlorophyll (Fletcher et al., 2000). PBZ-treated plants show an increase in cytokines and polyamines, delaying senescence produced by the synthesis of ethylene and abscisic acid. Likewise, studies by Hunter & Proctor (1992) on vine and blueberries, as well as Basiouny (1993) showed a delay in senescence based on PBZ treatments.
Some phyto-regulators used in fruit trees are explained in table (1), which shows the classification of phytohormones involved in the forced production of fruit trees, according to the literature cited in this document, as well as a growth bioregulator (paclobutrazol).
Table 1. Physiological mechanisms of action of phytohormones and growth bioregulators
| Classification | Examples | Mechanisms of action | Source |
| Auxins | Naphthaleneacetic acid, indolacetic acid | Two biosynthesis routes, the main route by means of tryptophan. They promote cell division and expansion, apical dominance, root development, stimulate gibberellin synthesis and indirectly stimulate stem growth. However, the ways in which these auxins are transported through the plant are very different. | (Fahad et al., 2015; Muñoz, 2016) |
| Giberellin | Giberellic acid | It is synthesized through the terpenoid pathway in the plastids and cytoplasm, derived from the isoprenoid biosynthetic pathway, and among its functions it stimulates cell division and elongation, and is related to stem growth, seed gemination, fruit development and flowering. | (Wang et al., 2020; Muñoz, 2016; Hernández-García et al., 2020) |
| Abscisic acid | Abscisic acid (ABA) | ABA synthesis occurs in chloroplasts via an indirect isopentenyl pyrophosphate (IPP) pathway, is associated with stress tolerance through stomatal closure, inhibits gibberellin inducible enzymes, stimulates root growth, seed dormancy, fruit ripening and abscission. | (Chávez et al., 2012; Muñoz, 2016) |
| Cytokinins | Zeatina and Thidiazhuron | It originates from the addition of isopentenyl pyrophosphate to N6, stimulates cell division and differentiation leading to the formation of roots, leaves, flowers and fruits, extends the vegetative period and helps in the accumulation of nutrients. | (Muñoz, 2016; Azcón-Bieto & Talón, 2008; Obregón, 2021; Nisler, 2018) |
| Polyamines | Putrescine, Spermidine, Spermine and Cadaverine | The synthesis of arginine in conjunction with S-adenosyl methionine, has been implicated in cell division and elongation, membrane stabilization, delay of senescence and reduction of the effect of water stress. | (Muñoz, 2016; Luna-Esquivel et al., 2014) |
| Triazole | Paclobutrazol | It is considered a growth retardant and inhibits gibberellin synthesis, decreasing competition between vegetative and generative growth, increasing flowering and carbohydrate translocation to the reproductive organ, improving stress tolerance and increasing chlorophyll and antioxidant enzyme content. | (López, 2015; Soumya et al., 2017) |
INCIDENCE OF PBZ IN OFF-SEASON CITRUS PRODUCTION
PBZ with its chemical structure [(2RS, 3RS)-1-(4-chlorophenyl)-4,4-dimethyl-2-(1H-1,2,4-triazole-1-il) pentan-3-ol] (Rademacher, 2016), belongs to the triazole group and is a synthetic growth retardant (Desta & Amare, 2021). By inhibiting the biosynthesis of gibberellins (Kumar et al., 2021), it blocks the oxidation reaction between kaurene acid and kaurenoic acid. This regulator aims to reduce or control the height of the stems and buds of plants (García et al., 2001), and acts on almost all plant species (Barrett, 2001).
Therefore, paclobutrazol has the dwarfed effect when used in citrus fruits, as it can reduce the excessive growth of buds, reason for which foliar applications should be made so as not to alter the normal growth of the roots, which favors the obtaining of an appropriately sized grafted plant (Garcia et al., 2001).
Gibberellic acid reduces flowering in citrus fruits, so it is necessary to apply inhibitory treatments of this hormone, such as PBZ. The application of regulators should be during induction or floral differentiation, since once this synthesis has been blocked there will be an increase in the concentration of starch in the roots, favoring flowering and the decrease of soluble sugars in leaves. In this way, the gibberellic acid/PBZ treatment program can be used to adjust the harvest (García et al., 2001).
On the other hand, PBZ increases the cold tolerance of citrus fruits, because it keeps the grafted buds in a dominant state during adverse conditions for the germination of buds. In addition, it is important to consider that this regulator is active at low concentrations applied by irrigation, causing a reduction in plant size by 20% (García et al., 2001).
PBZ is then considered to be a plant growth inhibitor in triazole class, and its main action is to inhibit gibberellic acid biosynthesis. By hindering vegetative growth, it will promote root growth, reduce the growth of stem and induce flower buds (Magdaleno et al., 2019). Its main effects include early flowering, early fruit maturity, restriction of vegetative buds, elimination of productive alternation, and increased production of perfect flowers and high yields (Cárdenas & Rojas, 2003).
Paclobutrazol has also been linked to the increase in chlorophyll content, acting on the oxidation of kaurene to kaurenoic acid, which are agents that cause toxicity risk, stimulating the production of flowers and giving way to the fruiting of the plant (Cázarez, 2015). For Quezada (2015), the application of gibberellin in doses of 0.35 g/L at a concentration of 10% in Washington Navel Orange trees (Citrus sinensis), allowed a yield of 54.171 oranges per hectare, while the witness that did not receive any treatment yielded 18.334 oranges per hectare, showing that regulators allow to increase the productive yield of citrus fruit trees.
PHYSIOLOGICAL EFFECT ON TRIAZOLE ACTION
Triazoles may be found under the following trade names Bonzi, Clipper, Cultar and Parsley (Fletcher et al., 1986). Its application can be foliar spraying or to the soil as drench (Rademacher, 2015) because triazoles can be transported in an acropetal (xylem) (Jung et al., 1985) and basipetal way, although they do not move too much in the phloem (Rademacher, 2000), which reduce the efficiency of foliar spraying because the PBZ will only act with the contact (Desta & Amare, 2021). Unlike applications of PBZ in drench that are more efficient at lower concentrations due to direct contact with roots, because it is where the greatest amount of gibberellic acid is synthesized and inhibited (Sopher et al., 1999), the useful life of the PBZ in the soil is 6 to 12 months depending on the edaphoclimatic conditions (Nørremark & Andersen, 1990)
One of the physiological effects of PBZ is that it inhibits the biosynthesis of gibberellic acid, blocking the oxidation of ent-kaurene to ent-kaurenoic acid (Rademacher, 2018). This process is performed through the deactivation of cytochrome-dependent oxygenase p450 (Rady & Gaballah, 2012), as noted in Figure (1) consequently, there is a decrease in cell division and elongation, causing a reduction in vegetative growth. The assimilates intended for this function will be used in generative growth, improving the uniformity of flowering (Sousa et al., 2020).
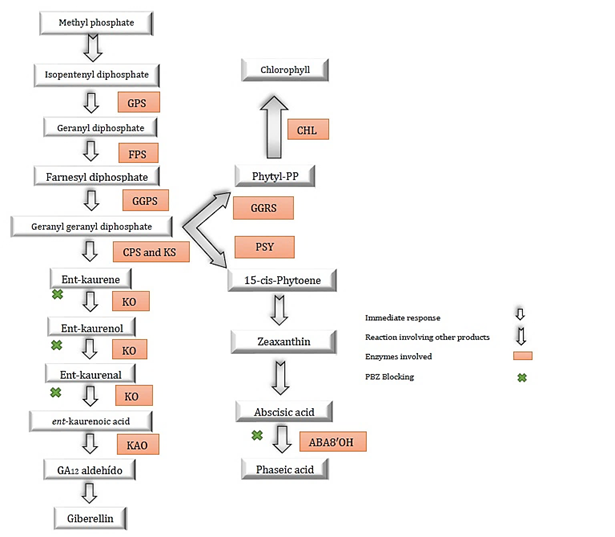
Figure 1. Terpenoid pathway for gibberellin biosynthesis, abscisic acid and chlorophyll formation points, in addition to areas blocked by enzyme inhibition due to paclobutrazol application. (Hedden & Sponsel, 2015; Soumya et al., 2017; Desta & Amare, 2021).
Likewise, it also affects the synthesis of abscisic acid because it is also synthesized by the terpenoid, and once the synthesis of gibberellin is blocked, an accumulation of precursors is formed, favoring the synthesis of abscisic acid (Rademacher, 1997). Increasing their concentrations in leaves, being their function the stomatic closure to prevent water losses through transpiration, decreases the growth of outbreaks and stimulates the growth of the roots for a greater water uptake (Desta & Amare, 2021).
SCIENTIFIC ADVANCES IN THE USE OF PBZ IN CITRUS PRODUCTION (ORANGE, LEMON AND TANGERINE)
The use of growth-retardant products in fruit trees has contributed to great advances in the study of citrus production, because it is a simple and effective strategy used to manage tree vigor, promote flowering and increase production (Rani et al., 2018).
Several studies have demonstrated the efficiency of PBZ to stimulate flowering in several citrus species such as C. aurantifolia (Snowball et al., 1994); C. sinensis (Acosta et al., 1994), among others. Studies carried out with the species C. aurantifolia proved it, where foliar applications of 1000 pp or to soil 15 mL a.i. L-1 of PBZ from April to August favored flowering between July and October, harvesting the fruits from December to March, where there is a productive deficit of the fruit, reaching the highest prices on the market (Medina et al., 1995). Similarly, Tripathi & Dhakal (2005) state that the application of 5 ml of PBZ dissolved in a liter of water applied in the tree neck area in July caused the induction of flowering in December, 70 days before the normal flowering date of the acid lime in Nepal.
Ariza et al. (2015) found that the use of phytohormones and biostimulators to stimulate flowering and improve the production and quality of Mexican winter lime reached a green color index with Ca+B+N SL (Biofol) and PBZ, and a yield of 9.763 kg/ha. Applying Ca+B+N SL to 1 L ha-1 the yield obtained with the application of paclobutrazol at 1,5 g m-2 was less than 50%, this coincided with the low yields reported in orange cv. Valencia when applying PBZ in Veracruz (Curti et al., 1990). However, Tripathi & Dhakal (2005) have a very important and critical process in the use of PBZ related to knowing how to manage times, because a treatment outside the growing season can produce different responses, being the most effective the early use of PBZ to induce flowering.
Phadung et al. (2011) in grapefruit experiments conducted in Thailand observed that when applying PBZ treatments at 750 mg/L twice a day every 15 days with water stress, i.e., suspension of water for a period of 12 days and then return to field capacity, it was observed that the treatment with water stress achieved significant flowering, obtaining the same results in the treatment with PBZ and water stress; with the difference that the last one did not show a noticeable decrease in the hydropower potential of leaves (Ψ). It could be related to the fact that PBZ stimulates the production of abscisic acid, and this in turn is linked to the stomatic closure to avoid water losses by perspiration, balancing the Ψ (Chaney, 2005), in addition to inducing flowering in grapefruit (Phadung et al., 2011).
An investigation on 'Tahiti' lime production (Citrus latifolia Tanaka), grafted with limonero 'Rangpur' (Citrus limonia Osbeck), subjected to water stress and applying PBZ obtained a higher number of flowers at concentrations of 762 mg per PBZ plants-1, yielding 192 flowers in every 100 knots, representing a 371% increase in plants subjected to water stress (Cruz et al., 2009).
Because of the latter, it is established that although citrus fruits have a high economic value due to their worldwide marketing, their production may be induced or inhibited by endogenous and exogenous factors. Exogenous factors that respond to floral induction include water stress, tropical, subtropical or temperate zones (Gravina, 2014). The endogenous factor affecting floral induction is the fruit load, due to the accumulation of gibberellin (Martinez-Fuentes et al., 2010) that reduces the CiFT genes (Muñoz et al., 2011), this effect of gibberellin was demonstrated by a study where they applied gibberellic acid at the inductive stage, reducing flowering by 72% compared to the witness; however, by applying PBZ they achieved opposite results by increasing flowering by 123% (Muñoz et al., 2012), hence its application becomes an important factor in fruit growing since it promotes off-season production so that a productive balance can be achieved to benefit producers and consumers.
A description (Table 2) of the main effects caused by PBZ on different fruit trees through foliar and soil applications was made.
Table 2. Efficiency of PBZ applied by edaphic and foliar application to different fruit trees
| Fruit | Form of application | Rate | Effect | Source |
| Mangifera indica. L | edaphic | 750 1000 mg l-1 | Shoot growth decreased by 30 and 13%, respectively. Both doses increased production compared to the control. The number of panicles and fruit quality increased. Increasing the dose of PBZ decreased TSS. | (Luo et al., 2016; Souza et al., 2016; Zhu & Stafne, 2019) |
| Mangifera indica. L | Foliar | 1000 2500 mg l-1 | Promoted 80% flowering and reduction of bud length. Increased flowering and bud differentiation by 60%. | (Barman & Mishra, 2018) |
| Persea americana Mill. | Edaphic Foliar Edaphic | 150 300 450 ppm/200L 1750 2500 mg l-1 | The treatments did not significantly influence shoot diameter. Increased shoot size reduction efficiency, reduced internode length. Increased fruit diameter, length and weight. Increased the percentage of flower buds per shoot by 35.32% with respect to the control with 12.60%, also obtaining a better flow of photo assimilates to the reproductive bud and higher yield. | (Brogio et al., 2018; Brioso & Layme, 2020; Ramírez, 2021) |
| Citrus aurantifolia | Zona del cuello | 5ml/1L | Induced flowering 70 days prior to normal flowering | (Tripathi & Dhakal, 2005) |
| Citrus paradisi | Foliar | 750 mg l-1 | It increased flowering in the PBZ treatments at 750 mg l-1 with the difference that the latter-maintained leaf water potential. | (Phadung et al., 2011) |
| Citrus latifolio Tanaka | 762 mg/ planta | Increased number of flowers, 192 flowers per 100 nodes in plants subjected to water stress. | (Cruz et al., 2009) | |
| Citrus clementina, citrus sinensis | Foliar | 2500 mg l-1 | Increased flowering relative to the control resulted in an increase in the number of nodes, inflorescences and a decrease in the number of vegetative buds. In leaves, the expression of CiFT and LFY genes, which are responsible for floral induction and differentiation, respectively, was increased. | (Munoz-Fambuena et al., 2013) |
CONCLUSIONS
Citrus are the consolidated fruit species with greatest economic importance worldwide. Its production is limited to the season of production and harvest, resulting in price reductions in the market, and affecting direct production costs.
Triazoles such as Paclobutrazol, which inhibits the synthesis of gibberellins and auxins, has been used in the search for management tools, slowing the growth of the plant by promoting the development of flowering and therefore achieving higher yields, being an important factor in inducing the plant to obtain an off-season crop and achieve increased yields.
According to the literature consulted, the importance of Paclobutrazol to induce out-of-season production of citrus fruits in several countries has been demonstrated; however, there is no data in Ecuador on the use of this compound for citrus production. Therefore, it is necessary to conduct new research that involves the productive process of citrus fruits with the use of paclobutrazol as a stimulant for the fruit yield of these plant species.
According to the analysis of the cited bibliography, it has been possible to force production, achieving a balance of prices in the market, so this is favorable for the producer.
In Ecuador, research is limited regarding PBZ, so it is vitally important to carry out research with PBZ in citrus, specifically in subtle lemon that is predominant in the province of Manabí and one of the problems focuses on the oversupply of production at certain times, so the results of these studies would become a direct support to the lemon producers, whose concern is the price of the product in the market. Antifungal properties are also attributed to PBZ, so by extending the research, results could be obtained on the type of control that would be achieved in relation to the type of fungal diseases in citrus.

