











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCCIÓN
La quinua es un cultivo ancestral del Perú que en la última década tuvo un incremento sostenido de la demanda nacional e internacional. Durante el periodo 2012-2019, el área cosechada se ha incrementado de 38 353 ha a 68 408 ha (MIDAGRI, 2021). La creciente demanda de dietas más saludables y funcionales a rescatado a la quinua, un alimento tradicional, con calidad nutricional superior a la de muchos cereales, por lo que fomentar su cultivo, es una estrategia que promueve la seguridad alimentaria principalmente para los países en vías de desarrollo (Schmidt et al., 2021). Las semillas sin gluten tienen un alto contenido de proteínas (16,3 g/100 g de masa) y un equilibrio excepcional de aminoácidos, con mayores contenidos de metionina, cisteína y principalmente de lisina (6 g/100g de proteína) (Burrieza et al., 2019; Filho et al., 2017; Pathan & Siddiqui, 2022), que es usualmente el aminoácido limitante en las dietas que excluyen la carne. Sin embargo, aún no se cuenta con información suficiente en cuanto a problemas fitosanitarios de este cultivo, principalmente para la región costa. Eurysacca melanocampta (Meyrick) y E. quinoae Povolny´ (Gelechiidae), con distribución en todo el Perú, han sido señaladas como las plagas más importantes en el cultivo de quinua (Rasmussen et al., 2003). Recientemente E. melanocampa, Macrosiphum euphorbiae Thomas, C., y Liriomyza huidobrensis Blanchard fueron señaladas como plagas importantes en la costa central (Lima) (Cruces et al., 2020). Otras plagas registradas en la costa peruana son Spodoptera frugiperda Smith & Abbot, S. eridania Stoll, S. ochrea Hampson, Chrysodeixis includens Walker, Myzus persicae Sulzer, Liorhyssus hyalinus (Fabricius), Nysius simulans Stål, Dagbertus spp., E. melanocampta Meyrick, Spoladea recurvalis Fabricius (Cruces et al., 2016)
Teniendo en cuenta la importancia de este cultivo en el Perú, es importante disponer de información sobre la presencia de insectos fitófagos de tal manera que nos permita optimizar el monitoreo e implementar estrategias de control oportuno de las especies plagas. Por ello, los objetivos fueron determinar las principales especies de insectos fitófagos y sus enemigos naturales en el cultivo de quinua, el porcentaje de infestación y el porcentaje de parasitoidismo en los insectos fitófagos.
MATERIALES Y MÉTODOS
Ubicación
Se empleó quinua de la variedad INIA 431-altiplano, en el campo experimental del Instituto Nacional de Innovación Agraria (INIA), en el distrito de La Molina, Lima, Perú (76,945224 W, 12,076648 S), desde el 21 junio a1 27 de septiembre de 2014. El área fue de 3432 m2 y limitaba por el noroeste con un canal de riego y un campo de palto con malezas (principalmente gramíneas), por el sudeste con una construcción, por el nordeste con un campo de quinua y por el sudoeste con un invernadero. Se realizó un manejo convencional con aplicación de plaguicidas, entre ellos clorpirifos y abamectina. El cultivo previo en esta área fue el frejol (Phaseolus vulgaris L.). Los datos de temperatura y humedad relativa fueron tomados de la estación meteorológica Alexander Von Humboldt de la Universidad Nacional Agraria La Molina (UNALM), Lima, Perú.
Metodología de evaluación
El campo fue dividido en 5 sectores (Figura 1), las evaluaciones se realizaron desde la emergencia hasta la cosecha, cada siete días, a partir de las 8:00 am, considerando un efecto borde de 1,5 m. Se emplearon las metodologías de, trampas de caída, evaluación por órgano de planta, red aérea y colecta de hojas minadas, propuestas por Sarmiento & Sánchez (2012).
Trampas de caída. Se empleó una trampa por sector (cinco trampas en total). Cada trampa consistió de un envase plástico de polipropileno (altura=11 cm, diámetro=8 cm), que contenía una mezcla de agua más 20 g de sal y cuatro ml de detergente líquido incoloro e inodoro.
Evaluación por órgano de planta. Se evaluaron cinco plantas por sector (25 plantas en total). En cada planta, se examinó el tallo, el brote principal, la panoja y dos hojas en la etapa de 4-6 hojas verdaderas, cuatro en la primera semana de panojamiento, seis en la segunda semana de panojamiento, dos al inicio de floración y primera semana de floración y cuatro desde la segunda semana de floración hasta grano maduro. En el caso de la evaluación de hojas, si estas se encontraban minadas, se trasladaron al laboratorio con el fin de determinar el porcentaje de infestación por larvas vivas.
Red aérea. - Se evaluaron dos surcos de 26 m por sector (10 surcos en total). A partir de la tercera semana (4-6 hojas verdaderas), se empleó una red de aro=30 cm Ø, longitud=80 cm. Los insectos capturados fueron sacrificados en cámara letal y luego trasladados en envases plásticos de polipropileno de 250 ml al laboratorio.
Colecta de hojas minadas. - Se colectaron 20 hojas por sector (total 100 hojas). Esta técnica se empleó para la evaluación de parasitoidismo en L. huidobrensis. Desde la emergencia hasta la etapa de 2 hojas verdaderas se extrajeron hojas minadas sin considerar su ubicación en la planta. En la primera semana de panojamiento las hojas fueron extraídas de la mitad inferior. A partir de la segunda semana de panojamiento, las hojas fueron extraídas del tercio medio. A inicio de floración y primera semana de floración, las hojas se extrajeron del tercio superior, esto debido a que la infección por mildiú eliminó las hojas de los tercios inferior y medio.
Manejo y procesamiento de muestras y datos provenientes de campo
Las muestras fueron examinadas en el Museo de Entomología Klaus Raven Büller (MEKRB) de la UNALM y registradas por separado por cada técnica de evaluación, separándolos por morfotipo y asignándoles un código a cada uno. Se registró el número total de individuos por morfotipo, por sector y fecha de evaluación en una hoja de cálculo. La mayoría de las muestras fueron preservadas en alcohol al 75% y un grupo fue sometido a montaje para su identificación.
Montaje
Los montajes en alfiler entomológico se realizaron de acuerdo con Timothy & Oseto (2020). Los Aphididae fueron montados en láminas siguiendo la técnica señalada por Blackman & Eastop (2000), Thripidae siguiendo la técnica de Mound & Kibby (2005) y para Braconidae (Aphidiinae) se empleó la metodología de Rosen & DeBach (1979).
Identificación
Para la identificación taxonómica se emplearon las siguientes claves: para Agromyzidae, Korytkowski (2014); para Cecidomyiidae, Gagné (1994); para Aphididae, Blackman & Eastop (2006); para Thysanoptera, Mound & Kibby (1998); para Crambidae, Munroe (1973); para Aphidiinae (Aphididae), Kavallieratos et al. (2010), Nazari et al. (2012) y Rakhshani et al. (2015); para Pteromalidae, De Santis (1987) y para Eulophidae, Peck (1963). Además, se realizó la comparación con los ejemplares del MEKRB.
Análisis de datos
Para determinar la cantidad de individuos registrados se realizó la sumatoria de los individuos colectados por cada fecha de evaluación, para cada técnica de evaluación. Se realizó el análisis de correlación entre las principales especies fitófagas y sus parasitoides, usando el coeficiente de correlación de Pearson.
RESULTADOS Y DISCUSIÓN
Insectos fitófagos asociados al cultivo de quinua
Se registraron 13 especies de insectos fitófagos, agrupados en cuatro órdenes (Thysanoptera, Hemiptera, Diptera y Lepidoptera) y 9 familias (Thripidae, Aphididae, Lygaeidae, Rhopalidae, Miridae, Cecidomyiidae, Agromyzidae, Noctuidae y Crambidae) (Tabla 1).
Enemigos naturales en el cultivo de quinua
Se registraron 29 morfoespecies de enemigos naturales, agrupados en cinco órdenes: Hemiptera, Neuroptera, Coleoptera, Diptera e Hymenoptera; y 15 familias: Berytidae, Miridae, Nabidae, Hemerobiidae, Chrysopidae, Dolichopodidae, Empididae, Syrphidae, Braconidae, Ichneumonidae, Pteromalidae, Eulophidae, Encyrtidae, Mymaridae (Tabla 2).
Tabla 1. Insectos fitófagos registrados mediante las distintas técnicas de evaluación en el cultivo de quinua en La Molina, Lima
| Orden/Familia | Especies | N° de individuos por técnica de evaluación | |||
| Órgano de planta | Red aérea | Trampa de caída | Colecta de hojas minadas | ||
| Thysanoptera | |||||
| Thripidae | Frankliniella occidentalis | 167 | 18 | ||
| Hemiptera | |||||
| Aphididae | Macrosiphum euphorbiae | 125 | 50 | 12 | |
| Myzus persicae | 89 | 33 | 13 | ||
| Lygaeidae | Nysius simulans | 2 | 3 | ||
| Rhopalidae | Liorhyssus hyalinus | 2 | 4 | ||
| Miridae | Dagbertus sp. | 10 | 2 | ||
| Diptera | |||||
| Agromyzidae | Liriomyza huidobrensis | 118 | 486 | 190 | |
| Cecidomyiidae | Prodiplosis longifila | 31 | 13 | ||
| Lepidoptera | |||||
| Noctuidae | Spodoptera frugiperda | 7 | |||
| Chloridea virescens | 2 | ||||
| Chrysodeixis includens | 2 | ||||
| Crambidae | Nomophila indistinctalis | 17 | 27 | ||
| Spoladea recurvalis | 4 | 3 | |||
Tabla 2. Enemigos naturales registrados mediante las distintas técnicas de evaluación en el cultivo de quinua en La Molina, Lima
| Orden/Familia | Morfoespecies | N° de individuos por técnica de evaluación | |||
| Órgano de planta | Red aérea | Trampa de caída | Colecta de hojas minadas | ||
| Hemiptera | |||||
| Berytidae | Metacanthus tenellus | 3 | |||
| Miridae | Rhinacloa pallidipes | 21 | 92 | ||
| Nabidae | Nabis capciformis. | 2 | |||
| Neuroptera | |||||
| Hemerobiidae | Hemerobius sp. | 6 | 4 | ||
| Chrysopidae | Chrysoperla externa | 16 | 12 | ||
| Coleoptera | |||||
| Carabidae | Blennidus peruvianus | 51 | |||
| Diptera | |||||
| Dolichopodidae | Chrysotus sp. A | 51 | 11 | ||
| Chrysotus sp. B | 271 | 43 | |||
| Empididae | Gen sp. A | 3 | |||
| Syrphidae | Allograpta exotica | 10 | |||
| Allograpta piurana | 3 | ||||
| Toxomerus sp. | 1 | ||||
| Hymenoptera | |||||
| Braconidae | Aphidius colemani | 60 | 208 | ||
| Bracon sp. | 3 | ||||
| Rogadinae sp. A | 1 | ||||
| Aphidiinae sp. A | 1 | ||||
| Ichneumonidae | Diplazon sp. | 2 | |||
| Campopleginae sp.A | 6 | ||||
| Neorhacodinae sp.A | 1 | 1 | |||
| Pteromalidae | Halticoptera arduine | 25 | 7 | 200 | |
| Gen sp.A | 7 | ||||
| Eulophidae | Diglyphus sp. A | 33 | 189 | ||
| Diglyphus sp. B | 5 | ||||
| Diglyphus sp. C | 2 | ||||
| Chrysocharis sp. A | 13 | 98 | |||
| Chrysocharis sp. B | 3 | ||||
| Closterocerus sp. | 5 | ||||
| Encyrtidae | Gen sp. A | 8 | |||
| Mimaridae | Gen sp. A | 3 | |||
Principales especies fitófagas y enemigos naturales registradas en el cultivo de quinua
Se consideraron importantes, debido al número de individuos registrados y por el impacto al cultivo, las siguientes.
Liriomyza huidobrensis (Diptera: Agromyzidae)
Las larvas fueron registradas a partir de dos hojas verdaderas. Los porcentajes más altos de infestación (porcentaje de hojas con larvas vivas) se registraron en las etapas de 4-6 hojas verdaderas (32%) y la segunda semana de panojamiento (15,3%). El porcentaje más alto de hojas minadas se registró en la etapa de 4-6 hojas verdaderas (40%). Al inicio de floración y primera semana de floración no se registraron larvas debido a una infección severa por mildiú, que ocasionó la caída de hojas en la planta. En la segunda semana de floración se volvió a registrar un 11% de infestación. A partir de entonces la población se mantuvo relativamente constante hasta la cosecha. Los adultos se registraron desde la primera evaluación por red aérea en la etapa de 4-6 hojas verdaderas hasta el final del cultivo. La población más alta de adultos (111 individuos mediante red aérea), fue registrada en la primera semana de floración.
L. huidobrensis se presentó durante toda la etapa de evaluación, similar a lo descrito en el cultivo de papa en la costa peruana (Cisneros & Mujica, 1998).
Las pérdidas económicas por la infestación de L. huidobrensis se han estimado en 15% en el cultivo de tomate (Lopes et al., 2019) y es capaz de infestar severamente diversas plantas en el Perú, en particular en localidades de la costa central (Weintraub et al., 2017). También se ha reportado una asociación de L. huidobrensis con el hongo Alternaria solani (Pleosporaceae) en cultivo de papa en Brasil (Soares et al., 2019).
Parasitoides recuperados de larvas de L. huidobrensis
Se recuperaron 502 individuos parasitoides de 1169 larvas. Los más abundantes fueron H. arduine (200 individuos, 39,84%), Diglyphus sp. A (189 individuos, 37,65%) y Chrysocharis sp. A (98 individuos, 19,52%). Otros parasitoides sumaron 15 individuos (3%). Estas especies fueron registradas durante toda la etapa de evaluación. H. arduine fue más abundante entre las etapas de 4-6 hojas verdaderas y panojamiento. Diglyphus sp. A fue más abundante en la etapa de grano lechoso. Hacia el final del cultivo la abundancia de parasitoides fue similar en las tres especies.
El primer registro de parasitoidismo ocurrió en la etapa de 4-6 hojas verdaderas (junio), con un porcentaje de parasitoidismo de 35,5%, luego 46,25% en la etapa de ramificación y 48,45% en panojamiento. Al inicio de la floración no se registraron parasitoides (debido a la ausencia de hojas con minas). En floración se registró 25% y fue incrementando hasta llegar a 55,88% en la etapa de grano maduro (septiembre, última evaluación) (Figura 2). Esto se asemeja a los resultados obtenidos por Cisneros & Mujica (1998) en el cultivo de papa, de 35% al inicio de la campaña en abril, hasta 55% de parasitoidismo al final del cultivo en agosto.
En el análisis de correlación entre las poblaciones de L. huidobrensis y los parasitoides se obtuvo un coeficiente de 0,966 (significativa en el nivel 0,01 (bilateral)) lo que indica una relación positiva muy fuerte entre ambas poblaciones (Hernández et al., 2014).
De siete especies de parasitoides registrados en el cultivo de quinua para la “mosca minadora”, seis corresponden a la familia Eulophidae y una especie a Pteromalidae, coincidiendo en cierta medida con lo que obtuvieron Cisneros & Mujica (1998) y Mujica & Kroschel (2013) en el cultivo de papa en Perú, y Callohuari et al. (2018) en el cultivo de tarwi, también en la costa peruana.
H. arduine fue más abundante en las primeras etapas de desarrollo del cultivo; a partir de floración, Diglyphus sp. A, fue más abundante. Estos resultados se asemejan a lo obtenido por Redolfi et al. (1985), Sánchez & Redolfi (1988), Cisneros & Mujica (1998), Mujica & Kroschel (2011) y Mujica & Kroschel (2013) en el cultivo de papa en la costa peruana, quienes señalaron que Diglyphus websteri (Crawford) y H. arduine fueron los más abundantes, por ende, los más importantes en la regulación de la población de “mosca minadora”.
Las condiciones de humedad relativa (76 - 96%) y temperatura (14,7 - 20,4 °C) durante este estudio fueron favorables para la presencia de L. huidobrensis (Mujica et al., 2017; Rodríguez-Castañeda et al., 2017).
Prodiplosis longifila Gagné (Diptera: Cecidomyiidae)
Solo se registraron larvas en brotes principales en la etapa de 4-6 hojas verdaderas (23 larvas en 25 brotes) y en ramificación (8 larvas en 25 brotes). Se registró un 20% de plantas infestadas en la etapa de 4-6 hojas verdaderas (4,6 larvas por brote en promedio) y 12% en ramificación (2,7 larvas por brote en promedio) (Figura 3). Los adultos solo fueron registrados mediante trampas de caída en bajo número.
Se conoce que P. longifila afecta todos los estados fenológicos que presentan brotes, en cultivos tales como, espárrago (Castillo et al., 2020), cítricos y tomate en Colombia (Duque-Gamboa et al., 2018; Velasco-Cuervo et al., 2016), sibidigua en Bolivia (Dhileepan et al., 2017), papa en Perú (Mujica & Kroschel, 2019) entre otros cultivos. Sin embargo, en este estudio no se registró infestación en las primeras etapas, pero sí después de la presencia de 4-6 hojas verdaderas hasta el inicio de panojamiento.
Luego de la aplicación de abamectina se registró un descenso en el número de plantas infestadas (Figura 3). La abamectina ha sido empleada para el control de P. longifila en papa en Perú (Mujica & Kroschel, 2019).
El rango de temperatura en el que se realizó este estudio fue de 14,7 °C a 20,4 °C, se encuentra dentro del rango favorable para el desarrollo de P. longifila (Castillo et al., 2020). Por otro lado, la humedad relativa estuvo en un rango de 76% y 96%, propicia para el desarrollo de P. longifila, como señalan Sarmiento (1997) citado por Valarezo et al. (2003) y Díaz (1981).
La larva produjo raspaduras en las hojas tiernas de los brotes principales al alimentarse, ocasionando como daño primario la deformación del brote y el cese del crecimiento de la planta. En algunos brotes afectados se desarrolló pudrición a causa de microorganismos patógenos, dadas las condiciones de humedad y deshechos de la alimentación de las larvas. Las plantas afectadas por esta especie desarrollaron una panoja deforme y menor en comparación a plantas no afectadas. Estas observaciones coinciden con lo señalado por Díaz (1981) para los cultivos de alfalfa, papa y pallar.
Myzus persicae, Macrosiphum euphorbiae (Hemiptera: Aphididae)
Myzus persicae fue registrado a partir de 4-6 hojas verdaderas (3 de individuos en 25 plantas), el mayor número de individuos se registró en las etapas de inicio de panojamiento (23 individuos) y segunda semana de panojamiento (19 individuos). Macrosiphum euphorbiae fue registrado a partir de dos hojas verdaderas (2 individuos en 25 plantas), sin embargo, el mayor registro de individuos fue en la segunda semana de grano maduro (70 individuos). El porcentaje más alto de plantas infestadas por áfidos (48%), se registró durante la segunda semana de panojamiento e inicio de floración. M. persicae fue más abundante entre las etapas de 4-6 hojas verdaderas e inicio de floración en comparación a M. euphorbiae, mientras que este último fue más abundante en la etapa de grano maduro.
M. persicae y M. euphorbiae son especies polífagas y cosmopolitas y han sido registrados en muchos otros cultivos tales como, fresa en Argentina (Cingolani & Greco, 2018), berenjena en Irán (Raeyat et al., 2021), tomate en Italia (Trotta et al., 2021), papa y tabaco en China (Liu et al., 2019; Zhou et al., 2021) donde además de perjudicar a las plantas por su alimentación, pueden convertirse en vectores de virus fitopatógenos.
Aphidius colemani (Hymenoptera: Braconidae)
Aphidius colemani fue registrado mediante red aérea durante todo el periodo del cultivo, sin embargo, en la evaluación por órgano de planta, solo fueron recuperados de Myzus persicae a partir de la etapa de ramificación. El mayor número de parasitoides recuperados fue en la primera semana de floración (15 individuos) lo que correspondió a un porcentaje de parasitoidismo de 83% (Figura 2).
Es probable que este parasitoide haya tenido una influencia importante sobre la población de áfidos, como ocurrió en el cultivo de trigo en Alemania donde las avispas parasitoides tuvieron el efecto más fuerte en la reducción del crecimiento de la población de áfidos en comparación a los predadores (M. H. Schmidt et al., 2003). En el cultivo de berenjena en Brasil, un parasitoide de la familia Braconidae fue uno de los factores que limitó el incremento de la población de áfidos (Leite et al., 2006).
El rango de temperatura en el que se desarrolló este estudio estuvo por debajo del rango de temperatura óptima para la actividad de A. colemani, que se sitúa entre 25 °C a 30 °C (Zamani et al., 2006).
Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae)
Se registró desde la etapa de 4-6 hojas verdaderas (2 individuos). En ramificación se registraron 10 individuos en 25 plantas, lo que representó un 16% de infestación (Figura 3). A partir de la floración, la población de esta especie se incrementó de manera sostenida, manteniendo ese patrón hasta la cosecha. En la etapa de grano maduro se registró el mayor número de individuos (49 individuos), con infestación de 72% (Figura 3). En la última evaluación se registró un descenso en comparación a la penúltima evaluación, esto podría deberse a que los adultos de F. occidentalis estarían migrando en busca de cultivos más jóvenes. Todos estos patrones de abundancia y movimiento se ajustan a lo descrito en estudios realizados en cultivos de lechuga, tomate y cucurbitáceas (Healey et al., 2017).
Uno de los factores más influyentes en la variación de la población de F. occidentalis probablemente fueron las etapas de desarrollo del cultivo. Las poblaciones altas ocurrieron a partir de grano lechoso; sin embargo, Atakan et al. (2016), Ren et al. (2020) y Healey et al. (2017) en otros cultivos, encontraron que la población máxima ocurrió en la etapa de floración. Lo ocurrido en el cultivo de quinua quizá se debió al estrés que sufrió la planta debido a la infección ocasionada por mildiu en la etapa de inicio de floración (defoliación total de la planta), en ese momento, el registro de trips fue muy escaso.
Las condiciones de temperatura y humedad relativa fueron favorables para el desarrollo de F. occidentalis. El umbral mínimo de temperatura para esta especie oscila entre 8-10 °C (Nielsen et al., 2021), mientras que las temperaturas entre 25 - 30 °C y sin fuertes variaciones, propician su desarrollo (Cao et al., 2019), permitiendo que ocurran varias generaciones en una sola temporada de cultivo (Reitz, 2009).
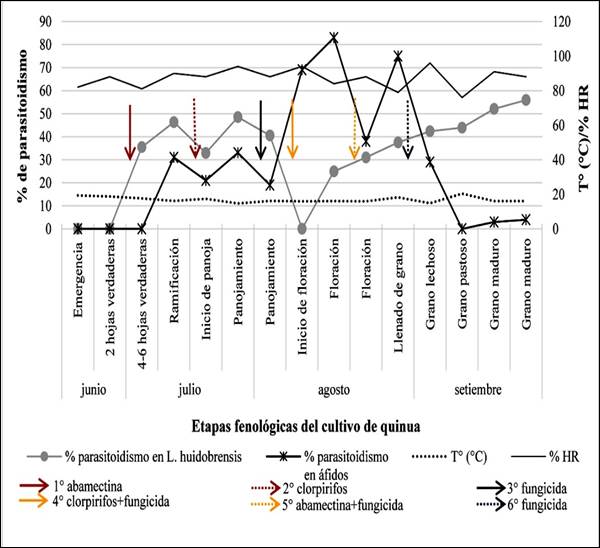
Figura 2. Porcentaje de parasitoidismo registrado en larvas de L. huidobrensis y áfidos en La Molina, Lima.

CONCLUSIONES
Las especies fitófagas más importantes registradas durante todo el desarrollo del cultivo de quinua son L. huidobrensis, P. longifila, M. persicae, M. euphorbiae y F. occidentalis. La infestación por larvas de L. huidobrensis es importante en las primeras etapas de desarrollo del cultivo, con una máxima infestación en la etapa de 4-6 hojas verdaderas (32%). P. longifila en estado de larva se presenta hasta la etapa de ramificación con máxima infestación de plantas (20%) en la etapa de 4-6 hojas verdaderas. M. persicae infesta principalmente brotes tiernos y M. euphorbiae principalmente panojas, con un 48 % de infestación por áfidos en panojamiento e inicios de floración. F. occidentalis registra mayor porcentaje de plantas infestadas en grano maduro (72%).
Los enemigos naturales más importantes son los parasitiodes A. colemani, H. arduine (Pteromalidae), Diglyphus sp. A y Chrysocharis sp. A (Eulophidae) y los predadores Chrysotus sp. B, Chrysotus sp. A, B. peruvianus, y R. pallidipes. Los parasitoides de larvas de L. huidobrensis alcanzan el porcentaje más alto de parasitoidismo en la etapa de grano maduro (55,88%). A. colemani muestra preferencia por M. persicae y se registra 84% de parasitoidismo en la etapa de floración.

