











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCCIÓN
Los manglares son ecosistemas que se distribuyen a lo largo de la línea costera, constituidos por árboles y arbustos que se desarrollan en zonas intermareales, con suelos salinos, inundados y anóxicos, constituyendo uno de los ecosistemas más productivos (Duke et al, 1998; Gómez, 2005). Son ambientes de gran importancia tanto ecológica como socioeconómicamente (Bouillon, 2008). Fungen como un filtro de nutrientes en las costas; además, realizan síntesis de materia orgánica obteniendo nutrientes inorgánicos provenientes de los ríos que ejerce un papel fundamental en la productividad de medio marino a los cuales están asociados (Pannier, 1979).
Los bosques de manglar se consideran uno de los sitios más apropiados para evaluar los efectos del cambio climático en las regiones tropicales, debido a la conexión que existe entre ellos (Pannier, 1992; Woodroffe, 1990). Estos se encuentran entre los ecosistemas más afectados por el cambio climático, especialmente por el aumento del nivel del mar, el viento, las olas, las corrientes y los patrones de tormentas (Yáñez et al. 1998).
Panamá posee una gran extensión costera, las cuales tienen condiciones muy favorables para el desarrollo de los bosques de manglar, siendo considerado uno de los países con mayor diversidad de manglares en el continente americano (ANAM-ARAP, 2013).
Los insectos herbívoros constituyen organismos reguladores de la productividad primaria, ejercen un papel fundamental en las interacciones tróficas y en el ciclaje de nutrientes de los bosques de manglar (McKee, 2002). Por esta razón, esta particularidad de los insectos se destaca la importancia de la herbivoría en los ecosistemas de manglares. Existes diversas investigaciones realizadas en otras regiones y que señalan la importancia y repercusiones ecológicas de los herbívoros sobre los ecosistemas de manglar (Feller, 1995, McKee, 2002; Lema Vélez et al. 2003, Romero et al. 2006; Tavares de Meneses & Peixoto, L. 2009).
En Panamá es muy escasa la información publicada sobre el papel de los insectos en los ecosistemas de manglar y específicamente en la región de la Bahía de Panamá. La poca información publicada y que trata sobre herbivoría es sobre el impacto de la herbivoría de larvas de Junonia genoveva (Cramer, 1780) (Lepidoptera: Nymphalidae) sobre las plántulas de Avicennia germinans L. (Lamiales: Acanthaceae) en la Bahía de Panamá, (Santos-Murgas et al. (2020). Por las razones antes expuestas establecimos el objetivo de identificar los principales insectos xilófagos que están atacando los árboles de mangles que se encuentran en el área protegida Refugio de Vida Silvestre sitio RAMSAR Humedal Bahía de Panamá.
MATERIAL Y MÉTODOS
Área de Estudio
Se realizaron observaciones biológicas y ecológicas en tres sitios, El embarcadero, Costa Sur y Don Bosco (Figura 1). En cada sitio se establecieron tres zonas u transepto de 100 metros de largo y 100 metros de ancho, desde el área de mareas hacia tierra firme, siendo la zona uno la que se encuentra más próxima a la línea costera, la zona dos que está en un punto intermedio entre el área de marea y el bosque, y la zona tres que se encuentra en la línea divisoria entre el manglar y el bosque (Figura 2). Los muestreos se realizaron cada 21 días durante cinco meses desde abril 2022 hasta agosto 2022. Está investigación contó con un esfuerzo muestreal de cinco meses, 150 días y 3600 horas.
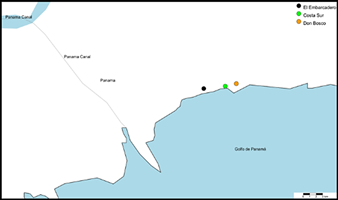
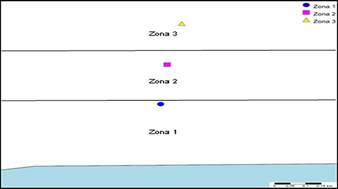
Figura 2 Esquema de la disposición de zonas de muestreo dentro de los bosques de manglar de la Bahía de Panamá.
El esquema de la Figura 2 describe la zonificación, donde la zona uno es la más próxima al mar, zona dos es intermedia y zona tres es la zona más pegada a los bosques tropicales próximos a los manglares.
Colecta de material biológico y trabajo de laboratorio
Se utilizaron trampas de intersección de vuelo para lo que se colocarán tres láminas (10 x 10 pulgadas) de plástico de color amarillo con pegamento (Figura 3A); las cuales fueron dispuestas en tres árboles de forma aleatoria en cada una de las zonas, a una altura de 1.60 metros del suelo; en total se colocaron nueve láminas con pegamentos por sitio de muestreo. Estas se mantuvieron durante 21 días, para luego ser recolectadas y transportadas al laboratorio para su separación e identificación.
El segundo método utilizado fue la colocación de sustratos compuestos de ramas de mangles para crías de escarabajos descortezadores, en cada una de las zonas de muestreo por sitios (Figura 3 B). Estas se dejaron colgados sobre arboles de mangles grupos en ocho ramas de una longitud aproximada de 15-20 pulgadas y un diámetro de 3-6 pulgadas, las cuales se amarraron en grupos y se dejaron colgadas al descubierto durante 40-50 días, para que fuesen colonizadas por escarabajos xilófagos. Pasado este tiempo fueron retiradas y transpor-tadas al laboratorio donde fueron colocadas en cajas de cría de mallas fina a una temperatura de 24 °C por un periodo de 15 - 25 días para la eclosión de los escarabajos y sus posibles parasitoides. Cada uno de los troncos fue revisado cuidadosamente y los individuos encontrados fueron colocados en viales de vidrio con alcohol al 70% para su posterior identificación.
El tercer método de colecta consta de trampas con atrayentes, las cuales fueron confeccionadas de manera artesanal, realizadas de botellas de plástico 2,5 L y se utilizó alcohol al 70% para preservarlos (Figura 3C), estas trampas fueron colocadas una en cada zonas o transeptos (tres por sitios) por un periodo de 21 días. Culminados este tiempo los especímenes fueron recogidos y colocados en viales de vidrio de 10 dm con alcohol al 70% los cuales fueron debidamente etiquetados con la información de colecta para ser transportados al laboratorio para su preservación e identificación.

Figura 3 Trampas utilizadas para la colecta de insectos xilófagos: A. Trampas de intersección de vuelo; B. Sustratos para crías de descortezadores; C. Trampas de botella con atrayentes.
El 4to método utilizado fue el de captura directa, realizando una búsqueda generalizada en cada uno de los transectos, duración aprox. de 60 min, en los cuales se colectaba insectos xilófagos de los árboles de manglar, los cuales se colocaron en viales 10 dm con alcohol al 70%, los cuales fueron debidamente etiquetados con la información de colecta para ser transportados al laboratorio para su preservación y posterior identificación.
Para la identificación de los insectos xilófagos asociados al bosque de manglar, se utilizó un estereoscopio Leica M205 y las claves taxonómicas propuestas por (Gasca, 2010; Pérez, 2021; Ratcliffe, 2011; Wodd, 1896).
Todo el material biológico se encuentra depositado en la Colección Nacional de Referencia del Museo de Invertebrados G. B. Fairchild, de la Universidad de Panamá (MIUP-UP).
RESULTADOS Y DISCUSIÓN
Las especies de mangle identificadas fueron Avicennia germinans, Laguncularia racemosa y Pelliciera rhizophorae. Donde está compuesta por Avicennia germinans siendo la especie que se presentó mayormente en las zonas uno y la zona do, mientras que en la zona tres si se pudo observar la presencia de las tres especies de mangle presente.
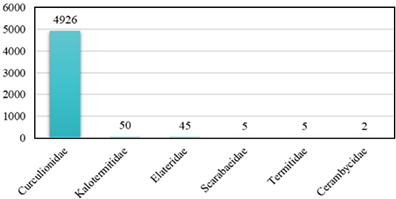
Se encontraron un total de 5033 individuos correspondientes a seis familias de insectos xilófagos asociados al bosque de manglar en los tres sitios de muestreo (Tabla 1). El sitio con mayor abundancia fue El Embarcadero con 3803 individuos seguido por Costa Sur con 710 individuos y Don Bosco con 520 individuos (Figura 4). La familia que presentó una mayor abundancia fue la familia Curculionidae con 4926 individuos que representa el 97% de los insectos xilófagos capturados, seguido por la familia Kalotermitidae con 50 individuos y Elateridae con 45 individuos. Las familias que presentaron una menor abundancia fueron, Scarabaeidae con cinco individuos, Termitidae con cinco individuos y Cerambycidae con dos individuos.
Tabla 1. Abundancia de familias de insectos xilófagos por sitio de muestreo
| Familias | Sitios | Total | ||
| Costa Sur | Don Bosco | Embarca-dero | ||
| Cerambycidae | 0 | 0 | 2 | 2 |
| Curculionidae | 678 | 480 | 3768 | 4926 |
| Elateridae | 16 | 11 | 18 | 45 |
| Kalotermitidae | 14 | 24 | 12 | 50 |
| Scarabaeidae | 2 | 0 | 3 | 5 |
| Termitidae | 0 | 5 | 0 | 5 |
| Total | 710 | 520 | 3803 | 5033 |
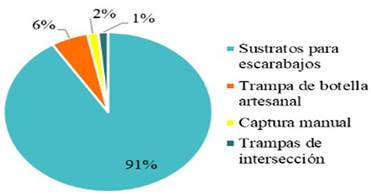
Evaluando el uso de distinto tipos de metodología obtuvimos que el método de colocación de sustratos para crías de escarabajos descortezadores o trampas de cebo fue el método que proporciono la mayor cantidad de individuos colectados con 4 584, la metodología de trampas con atrayentes capturo un total de 288 individuos, de los cueles el 100% de los individuos colectados pertenecen a la familia Curculionidae, las trampas de intersección de vuelo capturaron 78 individuos con la presencia de 3 familias (Curculionidae, Kalotermitidae y Elateridae) siendo la familia Elateridae la que presentó la mayor cantidad de individuos 35 colectados; y por método de colecta manual se capturaron un total de 85 individuos, en el cual se capturaron cinco familias de las seis encontradas durante el muestreo.
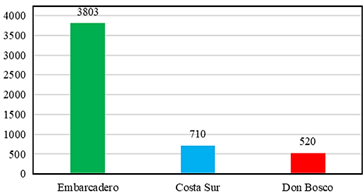
Figura 5 Abundancia de insectos xilófagos por zona de muestreo en los diferentes bosques de manglar.
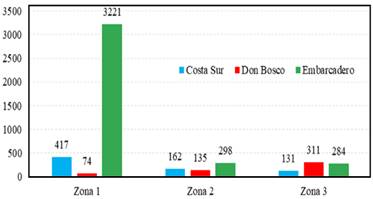
El sitio Embarcadero fue el que presento una mayor abundancia con 3803 individuos de cinco familias, siendo la familia Curculionidae (Coleoptera) la más abundante en este sitio, el método de captura con las trampas de cebo fue el más efectivo para la captura de individuos de la familia Curculionidae, obteniendo un 98% de los individuos de esta familia. De las tres zona muestreada, la zona uno fue la que presentó mayor abundancia de individuos y mayor riqueza de familias con 3221 individuos y cinco familias presentes, siendo las familia Curculionidae fue la más dominante sobre todas, con una abundancia de 3212 individuos, la zona dos presentó una abundancia de 298 individuos y dos familias, siendo las familia Curculionidae fue la más dominante con una abundancia de 289 individuos y la zona tres presenóo una abundancia de 284 individuos y tres familias siendo la familia Curculionidae dominante con una abundancia de 267 individuos (Figura 5, 7).
En el bosque de manglar ubicado en sitio Costa Sur obtuvimos una abundancia de 710 individuos y cuatro familias de insectos xilófagos, para los cuales la familia Curculionidae fue la más abundante con 678 individuos. El mayor número de individuos fue colectado en las trampas con atrayentes (alcohol al 70%) la cual capturo un total de 247 individuos pertenecientes al género Euplatypus, un género de barrenadores pertenecientes a la subfamilia Platypodinae. Para las zonas muestreadas, la zona uno fue la que presentó mayor abundancia con 417 individuos, seguida por la zona dos con 162 individuos y la zona tres con 131 individuos, la riqueza de familias tanto la zona uno, como la zona dos, fue de tres familias (Curculionidae, Elateridae y Kaloter-mitidae), mientras que en la zona tres se presentaron tres familias con la presencia de las familias Scarabaeidae, Curculionidae y Elateridae; con la ausencia de la familia Kalotermitidae.
La abundancia obtenida para el sitio Don Bosco fue de 520 individuos en cuatro familias de insectos xilófagos, de la cuales la familia Curculionidae fue la más abundante con 480 individuos. La mayor cantidad de individuos fue capturada con las trampas cebo, la cual fue más efectivo para la captura de individuos de la familia Curculionidae. La zona tres fue la que presentó una mayor cantidad de individuos con 311, mientras que la zona uno y la zona dos presentaron una abundancia de 74 individuos y 135 individuos respectiva-mente; siendo la familia Curculionidae la más abundante en todas las zonas (Figura 4).
Los 5033 individuos correspondientes a las seis familias de insectos xilófagos asociados a los bosques de manglar, del área protegida refugio de vida silvestre sitio RAMSAR, humedal Bahía de Panamá, contribuyen significativamente a la estructura trófica de los bosques de manglar de la Bahía de Panamá; esta explicación tiene su base en las transformaciones de materia orgánica originada por la producción de hojarasca y descomposición de la madera de los árboles de mangle efectuada por los insectos que habitan en el manglar (Prahl et al. 1990).
El embarcadero resultó ser el sitio que presenta la mayor cantidad de insectos xilófagos con 3803 individuos, siendo la zona uno en el sitio El Embarcadero y el sitio Costa Sur las que resultaron con un mayor número de insectos xilófagos y herbívoros, mayoritarmente de la familia Curculionidae. Este material vegetal consumido por los herbívoros va a depender de diferentes factores físicos o procesos biológicos que se estén dando en los ecosistemas del manglar. De esta manera, la acción de estos insectos puede regular la productividad primaria y ejercer un papel fundamental en las interacciones tróficas de los bosques de manglar (Robertson 1991).
Tabla 2 Insectos xilófagos colectados en los bosques del manglar de la Bahía de Panamá
| Especies | Nº de individuos |
|---|---|
| Aeolussp. 1 | 9 |
| Aeolussp. 2 | 8 |
| Aeolussp. 3 | 12 |
| Aeolussp. 4 | 1 |
| Aeolussp. 5 | 4 |
| Aeolussp. 6 | 1 |
| Aeolussp. 7 | 3 |
| Aeolussp. 8 | 1 |
| Chalcolepidius sp | 5 |
| Conotrachelus sp. 1 | 32 |
| Cryptotermes sp | 9 |
| Euplatypus Parallelus | 294 |
| Incisitermes sp | 41 |
| Lacon palliatus | 1 |
| Mallodon sp | 2 |
| Nasutitermes sp | 5 |
| Phileurus sp | 5 |
| Xyleborus ferrugineus | 821 |
| Xyleborus sp. 3 | 10 |
| Xyleborus sparsipilosus | 3769 |
| Total | 5033 |
Esta investigación contribuye a explicar la función de los insectos xilófagos en los bosques de manglar. Investigaciones realizadas anteriormente han demostrado que factores como el efecto de los nutrientes, la distancia al continente y la intervención humana pueden incrementar la tasa de consumo por herbívoros y xilófagos (Onuf et al., 1977; Johnstone, 1981; Lacerda et al., 1986; Farnsworth y Ellison, 1991; Blanchard y Prado, 1995; Romero, et. al., 2006). También hay que mencionar que pocos trabajos han tratado de establecer las características propias que realiza cada especie de insectos herbívoros sobre el manglar, como por ejemplo sus diferencias y el tipo de destrucción que causan sobre el material vegetal. Otro tema por destacar es que la mayoría de las investigaciones que se han realizado sobre esta temática se ubican principalmente en el Caribe de América y en ambientes diferentes a los estuarios (Onuf et al. 1977, Johnstone 1981, Lacerda et al. 1986, Robertson y Duke 1987, Farnsworth y Ellison 1991, Anderson y Lee 1995, Feller 1995, Stowe 1995; Romero, et. al. 2006).
Las diferentes metodologías utilizadas en el muestreo arrojaron resultados poco similares, pudiéndose considerar las de trampa de cebo y las de trayentes bastante eficientes en la captura de insectos de la familia Curculionidae, ya que capturan la mayor parte de insectos en todo el muestreo; nuestros resultados coinciden con los obtenidos por Kovach and Gorsuch (1985) y Flores, R. et. al. (2014), que reporta el uso de la trampa multidireccional, para la captura de insectos de la familia Scolytidae; utilizando como atrayente etanol, a una concentración del 70%. Este resultado nos indica que es un método de captura eficiente y eficaz, que puede utilizarse como complemento al método de recolecta manual o directo, en este tipo de investigaciones donde se haga uso del estudio de especies claves en ecosistemas de manglares Romero, et. al. (2006). Las trampas de intersección de vuelo resultaron ser un poco más generalista pero no tan eficientes, y el método de captura directa es el que aporta la mayor riqueza de familias y abundancia de cada una de las familias se xilófagos asociados al bosque de manglar.
La familia de insectos xilófagos que se encontró asociadas al bosque de manglar y presento la mayor abundancia fue la familia Curculionidae, siendo la subfamilia Scolytinae y Platypodinae las de mayor importancia de este estudio, sanos (Ploetz et al., 2013), encontró que algunas especies de Platypodinae atacan árboles debilitados, incluso algunas especies que pueden colonizar y hasta causar la muerte a árboles sanos que coinciden con nuestros resultados, ya que se evidencio la presencia de muchos individuos en ramas y troncos secos.
Los miembros de la subfamilia Scolytinae, muchos son considerados plagas graves para muchas especies de árboles, estos se han registrado afectando arboles de mangle en diferentes países (Castillo, 2001; Martínez et al., 2017; Rabinowitz,1977).
Para la subfamilia Scolytinae encontramos 4584 individuos, siendo la zona uno del El Embarcadero el sitio que presentó un mayor número de individuos, colocándolo como el sitio más vulnerable de todo el muestreo. Tanto El Embarcadero como en Costa Sur la zona uno resultó el sitio que se observó la mayor cantidad de insectos xilófagos, por lo cual podemos presumir que las zonas más próximas al mar es la que será más propensa a sufrir ataque por insectos xilófagos, ya sea por alteraciones en la salinidad, cambios climáticos y fenómenos como el niño y la niña.
En el sitio Don Bosco la zona tres fue la que presentó una mayor suma de insectos xilófagos manteniéndose por debajo de lo encontrado en El Embarcadero y Costa Sur; para Don Bosco la cantidad de insecto encontrados fue menor que la otros sitios, por ende podemos suponer que los árboles de este sitio se mantiene en condiciones apropiadas para la defensa contra estos insecto xilófagos que se aprovechan de árboles enfermos o en condiciones de estrés, aunque la zona tres tuvo una mayor cantidad de insecto en comparación con los demás sitios, esto se puede deber a que se encuentra en contacto con el bosque primario que posee otro tipo de vegetación.
La zona uno y la zona dos de El Embarcadero y Costa Sur, se notó una alta cantidad de árboles con la presencia de ataques sobre la corteza realizados por insectos de la subfamilia Platypodinae (Figura 8), pertenecientes al género Euplatypus, siendo sugerido por Wood & Bright (1992) como uno de los géneros de Platypodinae más destructivos y con una amplia distribución mundial, los cuales no poseen una preferencia especifica por alguna especie de árboles. La especie fue identificada como Euplatypus parallelus, las cual construyen galerías que suelen llegar hasta el duramen de la planta, las cuales pueden ser causante de la muerte de alguno arboles débiles.
Es probable que una variedad de factores hayan sido los responsables de las diferencias del consumo del material vegetal por parte de los insectos xilófagos observadas entre las especies del manglar y las zonas muestreadas. Entre los factores pudiéramos mencionar la salinidad, la intervención de las mareas, incluso la influencia de factores antropogénicos contaminantes.
Es importante mencionar que las condiciones relativas de los factores ambientales, la condición del bosque, el grado de intervención de las zonas del manglar donde se encuentran los árboles, son los principales factores que influyen sobre el consumo de la materia vegetal, por parte de los insectos xilófagos sobre todas las especies de mangles estudiadas; además, pueden determinan la tasa y el tipo de consumo que sufren las especies de mangle a lo largo del estuario.

Figura 8 Daño caudado por Euplatypus parallelus (Platipodinae) sobre Avicennia germinans. A. Daño inicial por E. parallelus. B. Distribución del daño causado por E. parallellus en la corteza de A. germinas. C. Orificios y canales realizados por E. parallellus en los tejidos interno del tronco de A. germinas. D. E. parallellus atrapado por la resina del mangle y evidencia del daño.
AGRADECIMIENTOS
Agradecemos al Instituto Smithsonian de Investigaciones Tropicales, Panamá y a la administración de proyecto “Impact of Insects Herbivory on Mangroves in the Ramsar Wetland Wildlife Protected Area, Juan Diaz, Bay of Panamá” por proveer los fondos para la realización de esta investigación. También, deseamos agradecer al Profesor Alfredo Lanuza G. por corroborar las identificaciones de los escarabajos xilófagos colectados en la Bahía de Panamá. .
CONCLUSIONES
El Embarcadero fue el sitio con mayor número de insectos xilófagos capturados, con 3.803 individuos. De las tres áreas muestreadas, las áreas más cercanas a la costa (Zona 1) El Embarcadero y Costa Sur presentaron la mayor cantidad de insectos xilófagos. Las familias de insectos xilófagos encontradas fueron Kalotermitidae, Termitidae, Cerambycidae, Curculionidae, Elateridae y Scarabaeidae. La familia de escarabajos Curculionidae fue la más comúnmente encontrada, siendo las subfamilias Scolitinae y Platypodinae las más importantes, por su impacto en las cortezas de los árboles. El 91% de todos los insectos xilófagos capturados pertenecían a la subfamilia Scolytinae, mientras que el 6% pertenecía a la subfamilia Platypodinae. Euplatypus parallelus, afectó en gran cantidad a individuos del mangle Avicennia germinans, ocasionando severos daños a los troncos de esta especie de mangle.

