











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCCIÓN
En general los animales utilizan señales acústicas para comunicarse, lo que implica que las propiedades de estas señales pueden estar sujetas a una fuerte selección. Se conoce también que la adaptación acústica predice que las especies en hábitats densos emiten sonidos de menor frecuencia que las de áreas abiertas porque los sonidos de baja frecuencia se propagan más lejos en la vegetación densa que los sonidos de alta frecuencia. La frecuencia de la señal también puede estar bajo selección sexual porque se correlaciona con el tamaño del cuerpo (Mikula et al., 2020).
En ese sentido para garantizar una comunicación acústica eficaz, las señales deben llegar a los receptores de la forma menos distorsionada posible. Los animales utilizan diversas estrategias a corto y largo plazo para evitar la degradación y el enmascaramiento de la señal (Budka, 2023).
Aunque muchas formas de comunicación animal no dependen del orden en que se combinan los componentes de las señales para que sean efectivas, existe evidencia de que el orden sí importa para algunos sistemas de comunicación (Campbell et al., 2023).
En cuanto a las aves emiten sonidos a partir de la siringe los cuales se llaman vocalizaciones, estas se clasifican en llamadas y cantos. En general, los llamados son sonidos de corta duración formados por una o pocas sílabas simples o medianas en complejidad. Los llamados son producidos tanto por machos como por hembras a lo largo del año, generalmente son innatos en todos los grupos de aves y normalmente pueden relacionarse con funciones específicas como vuelo, amenaza, alarma, forrajeo o contacto con parientes u otros miembros de la misma especie, hasta ahora se sabe que los cantos son aprendidos solamente en tres grupos de aves: aves canoras(oscinos), pericos y colibríes. Para poder estudiar los cantos en las aves y cualquier otra señal acústica, es necesario medirlos, esto se hace a partir de los espectro-gramas, que son representaciones visuales del sonido. Las características o medidas más usadas son la frecuencia mínima, frecuencia máxima, frecuencia pico y ancho debanda, las cuales miden la variación en el tono de los cantos, las características temporales más usadas son la duración del canto, el tiempo entre cantos o entre sílabas y tasa de canto. Las propiedades del sonido de los cantos de las aves nos ayudan a identificar a las especies, incluso ciertas propiedades pueden corresponder a determinados grupos de aves. (Moran & Gonzales, 2022).
Las vocalizaciones que producen las aves son una herramienta que ha sido ampliamente usada, por su papel en la delimitación e identificación de especies. En ese sentido representan un carácter diagnóstico que facilita la discriminación de entidades taxonómicas, esto ha sido de gran utilidad en la confirmación y registro de especies que dados sus comportamientos y requerimientos de hábitat son de difícil observación (Fernández et al., 2023).
De acuerdo con Griffiths & Aaronson (2023), la variación en la vocalización de especies del género Falco puede ser sólo el resultado de la variación en la estructura anatómica, siendo el tamaño el factor principal. Los autores encontraron que cuanto más grande es el ave, menor es la frecuencia máxima.
Por otro lado, las técnicas de minería de datos provienen de la inteligencia artificial y de la estadística, dichas técnicas son plasmadas en algoritmos, que después se aplican sobre un conjunto de datos para obtener resultados (Peralta et al., 2022). Dentro de la minería de datos se usa el análisis clúster el cual engloba una serie de técnicas que pueden ser usadas para crear agrupamientos, más concretamente un método clúster es un proce-dimiento estadístico multivariante que comienza con un conjunto de observaciones y los reorganiza en grupos o clúster con las siguientes cara-cterísticas: a) cada grupo o clúster es homogéneo o compacto con respecto a ciertas características, es decir las observaciones en cada grupo son similares entre sí,b) cada clúster ha de ser diferente de los otros con respecto a las mismas características, es decir las observaciones de un grupo serán diferentes de las observaciones de los otros grupos y estas diferencias serán mayores cuantos más alejados estén los clústers entre sí. Parte de los anális clúster son los llamados métodos jerár-quicos; los cuales se caracterizan fundamen-talmente porque tratan de agrupar varios clústers para formar uno nuevo o bien para dar origen a otros dos partiendo de uno ya existente, de tal forma que, si sucesivamente se va realizando este proceso de aglomeración o división, se minimicen las distancias o se maximicen similitudes entre los grupos (Broncano de Pablo, 2018).
En ese sentido el objetivo de este trabajo fue conocer que especies de las familias Falconidae y Strigidae guardan similaridad entre si al pertenecer a un mismo grupo después de aplicar algoritmos de aprendizaje no supervisado (análisis clúster), tomando como base las características cuantitativas de sus vocalizaciones.
MATERIAL Y MÉTODOS
Se descargaron grabaciones de la base de datos de acceso abierto Xeno canto de las especies: Falco sparverius, Falco rufigularis, Falco femoralis, Falco peregrinus, Falco columbarius, (Familia Falconidae), Glaucidium peruanum, Glaucidium jardinii, Glaucidium hardyi, Megascops roboratus, Aegolius harrisii y Glaucidium brasilianum (Familia Strigidae).Debido a que hubo diferente cantidad de grabaciones de cada una de las especies y para poder analizar igual cantidad de grabaciones se realizó un muestreo no probabilístico por conveniencia de 53 audios, se eligió esta cantidad ya que la especie Megascops roboratus es la que tiene la menor cantidad de audios los cuales son 53.
Una vez descargados las grabaciones, desde el 3 de enero al 3 de diciembre del 2022 con ayuda del programa Audacity se limpiaron los audios de cualquier ruido de fondo con la opción efecto EQ y filtros y ecualizador gráfico. Una vez limpios los audios se obtuvieron datos de frecuencia mínima, media y máxima, duración mínima, media y máxima, amplitud mínima, media y máxima, detección de picos de frecuencia, factor cresta, índice de complejidad acústica y mediana de envolvente de amplitud. Estos se calcularon con la ayuda de la librería Seewave (Sueur et al., 2008) del programa R y el programa Raven pro Lite y se colectaron en una matriz de datos. En esta matriz de datos las columnas estuvieron formadas por las variables: frecuencia mínima, media y máxima, duración mínima, media y máxima, amplitud mínima, media y máxima, picos de frecuencia, factor cresta, índice de complejidad acústica y mediana de envolvente de amplitud. Y las filas por las especies estudiadas.
Con la matriz de datos, en el programa R se realizó un arreglo para que el nombre de las especies el programa no lo lea como una variable cualitativa. Para lo cual se reordenó el nombre de las especies
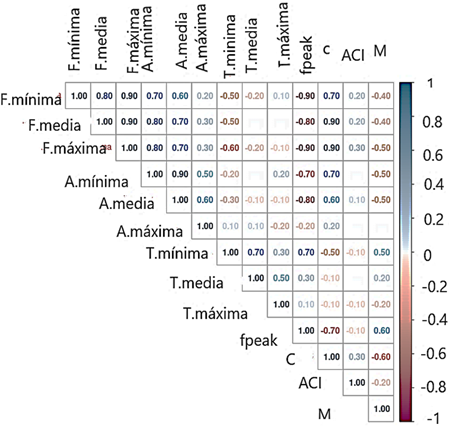
en una columna aparte, para que de esa manera solo queden las variables cuantitativas. Esto se realizó con la ayuda de la librería textshape (Rinker, 2021). Luego se prepararon los datos a través de un escalamiento. Asimismo, se midió la correlación entre las variables con la librería PerformanceAnalytics (Peterson & Carl, 2020) de R. Posteriormente se evaluó la tendencia de los datos al agrupamiento de acuerdo con el esta-dístico de Hopkins, con la ayuda del paquete Hopkins (Wright, 2022) del programa R, el cual establece que valores entre 0 - 0,3 indican datos espaciados regularmente, datos alrededor de 0,5 indican datos aleatorios y valores entre 0,7 - 1 indican datos agrupados. Luego con los algoritmos: kmeans, PAM, CLARA y clúster jerárquico, y con la ayuda del paquete clValid (Brock et al., 2008) de R se eligió el más adecuado, utilizando para este fin los índices de conectividad, de Dunn y la silueta teniendo en cuenta que el algoritmo más adecuado para aplicar a los datos debe minimizar el primer índice y maximizar los últimos.
Con las librerías clúster (Maechler et al., 2022) y facto extra (Kassambara & Mundt, 2020) se procedió a aplicar el algoritmo más idóneo, que para estos datos fue: el clúster jerárquico, específicamente el aglomerativo y se comparó cual, entre los métodos single, complete y Ward era el más idóneo, usando para ello el coeficiente de aglomeración, teniendo en cuenta que el coeficiente más cercano a 1 es el que mejor agrupa a las observaciones. Una vez obtenido el mejor método de clúster jerárquico aglomerativo, se procedió a graficarlos. Todo esto se realizó con las funciones hclust y agnes de la librería clúster.
RESULTADOS Y DISCUSIÓN
De acuerdo con la Tabla 1, cada especie y cada familia presenta sus características específicas de frecuencia, amplitud y duración. De acuerdo con Moran & Gonzales (2022), las propiedades del sonido de los cantos de las aves nos ayudan a identificar las especies, incluso ciertas propiedades pueden corresponder a determinados grupos de aves. Por ejemplo, las palomas (familia Columbidae) emiten cantos de bajas frecuencias (graves), con sílabas largas, simples (sin modulaciones de frecuencia) y de baja intensidad. Los saltaparedes (familia Troglodytidae) emiten cantos muy diversos, pero algunas propiedades que los caracterizan son trinos y sílabas con modulaciones de frecuencia en un rango aproximado de 1 a 10 kHz, según sea la especie y generalmente son intensos. Otros trabajos también corroboran lo mencionado, por ejemplo, López (2015), quien encontró diversas frecuencias, amplitudes y duraciones en los cantos de los distintos taxa del género Bubo en América.
Esta diferencia de las características de las vocalizaciones no solo se aprecia entre las aves sino también en otros grupos como anfibios (Batallas & Brito, 2016) o en los quirópteros (Malo de la Molina, 2011). Las duraciones, amplitudes y frecuencias también están relacionados con el comportamiento de la especie a estudiar.
Orellana (2014) encontró diversas voces del zorzal andino (Turdus chiguanco) cada voz asociada a un comportamiento y a una frecuencia, amplitud y duración característica.
Tabla 1 Frecuencia(f), amplitud(A), duración(T), Picos de frecuencia(fpeak), factor cresta(C), índice de complejidad acústica (ACI) y mediana de envolvente de amplitud(M) de las vocalizaciones de las especies estudiadas
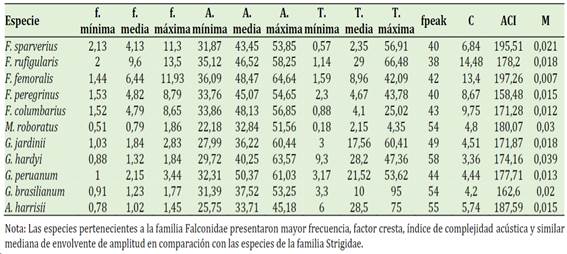
En la Tabla 1 también podemos observar que unas especies tienen frecuencias más altas que otras. De acuerdo con Arévalo (2019), estos cambios se deben al hábitat donde se encuentra el ave, así aquellas que viven en zonas urbanas presentan una frecuencia más alta esto se pudo apreciar por ejemplo en Falco sparverius y Falco rufigularis quienes viven en zonas urbanas, además de las zonas donde viven estos animales también influye la masa corporal la cual está relacionada con las frecuencias mínimas de canto. Wheeldon et al. (2022), sin embargo, mencionan que los cantos de Laniarius atroflavus se propagan mejor en los bosques que en sitios perturbados cubiertos de arbustos y los tipos de canciones masculinas utilizadas para el canto transmitido se propagan más lejos que las canciones femeninas. Por otro lado, Miluka et al. (2020) encontraron en un estudio global de paseriformes que la frecuencia de las canciones disminuye con el aumento de la masa corporal y con el dimorfismo de tamaño sexual sesgado por los machos; mas no encontró relación prevista entre frecuencia y hábitat.
De acuerdo con la Tabla 1 existen más picos de frecuencia en la familia Strigidae, de acuerdo con Loayza (2021) esta métrica funciona bajo el supuesto de que a mayor cantidad de picos de frecuencia existe una mayor actividad acústica.
En la Figura 1 el factor cresta (C) se relaciona con las frecuencias mínimas (0,70), máximas y medias (0,90) positivamente, También podemos observar que las frecuencias, duraciones y amplitudes mínimas y medias tienen relaciones positivas entre ellas. El factor cresta mide cuanto fluctúa la amplitud y por lo tanto podemos mencionar que en los Falconiformes la amplitud de la señal varía mucho o lo que se traduciría de acuerdo Morán & Gonzales (2022) que el volumen de la señal varía. Por otro lado, este factor al guardar una correlación positiva con la frecuencia medía y máxima nos da la idea que estaría relacionado con tonos de las vocalizaciones.
El índice de complejidad acústica (ACI) es mayor en unas especies más que en otras y en la familia Falconidae más que en la Strigidae según Lozano et al. (2014) al analizar los ACI, pero a nivel de especies no de comunidad mencionan que el mayor ACI por especie puede deberse al repertorio acústico que es muy variado. Por otro lado, Pieretti et al. (2011) encontraron el ACI relacionado con la cantidad de notas de las vocalizaciones (correlación de Spearman = 0,94) más que de las frecuencias. En este estudio tampoco se encontraron correlaciones del ACI con las frecuencias.
En este trabajo el estadístico de Hopkins resultó ser de 0,90 lo que indica que esta data es clusterizable. La mayoría de los índices indican que el mejor algoritmo para los datos es la agrupación jerárquica (Tabla 2).
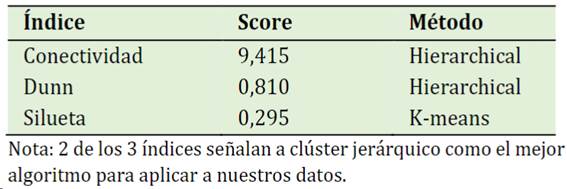
Además, en el algoritmo de clúster jerárquico el método que presentó mayor coeficiente de aglomeración fue el Ward (0,67), mientras que el método single y completo sus coeficientes fueron de: 0,284 y 0,582 respectivamente. Por lo que se aplicó el primer método.
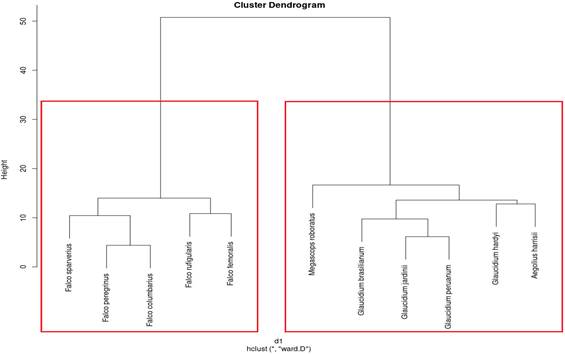
De acuerdo con la Figura 2, a partir de las características de sus vocalizaciones las 11 especies estudiadas se juntaron con miembros de su propia familia si lo comparamos con la lista de aves del Perú (Plenge, 2023). De acuerdo con Moreno (2006), las vocalizaciones permiten la comunicación de las aves; asimismo Vásquez & Vásquez (2018) señalan también que el canto permite la comunicación de individuos de la misma especie o entre especies distintas, buscando también modificar el compor-tamiento del individuo que la recibe, por ejemplo, Campbell et al. (2022) encontraron que individuos de menos un año de Poecile atricapillus son capaces de percibir y actuar según categorías de llamadas. Otro ejemplo de cómo el canto influye en el comportamiento lo señala Romani et al. (2023) quienes encontraron que los adultos de Ardea cinerea utilizan una amplia gama de tipos de llamadas, principalmente relacionadas con el cortejo y la fase reproductiva. Las vocalizaciones son emitidas especialmente por los machos y son un elemento clave durante la atracción de pareja, la construcción de nidos, en los eventos de reproducción y en el fortalecimiento de los vínculos dentro de las parejas. Por lo que agrupación resultante producto de la similaridad de sus vocalizaciones sería de alguna manera para asegurar su supervivencia en la naturaleza. Por otro lado, Vásquez & Vásquez (2018) también encontraron similitud en las vocalizaciones de las especies de diferentes familias las cuales fueron Mimus longicaudatus (Mimidae) y Campylorhynchus fasciatus (Troglodytidae) en especial en sus frecuencias máximas y mínimas.
En la Figura 2, las especies de la familia falconidae se separan de la Strigidae, esto se podría explicar de acuerdo a Batallas & Brito(2016) a condiciones ambientales y de contexto social. Calderón (2022) concluyó que el factor morfológico es relevante tanto por la producción de nuevas vocalizaciones aumentando y disminuyendo los repertorios en cinco especies de la familia Rhinocryptidae, incluso estaría afectado a los rangos de actividad junto a los aspectos reproductivos de estas aves debido a los costos energéticos que demanda la reproducción que no se pueden sobreponer con las vocaliza-ciones. En 7 especies del genero Falco, Griffiths & Aaronson (2023) encontraron que la variación en la vocalización de estas especies puede ser sólo el resultado de la variación en la estructura anató-mica, siendo el tamaño el factor principal. Los autores encontraron que cuanto más grande es el ave menor es la frecuencia fundamental y más lenta es la tasa de cambio de frecuencia (pendiente de frecuencia más pequeña) dentro de las notas, también encontraron una fuerte correlación entre la pendiente de frecuencia y de las estructuras de la siringe. Vargas (2021) evaluó la comunidad de aves con relación al paisaje sonoro como una propuesta para el establecimiento de un Corredor Biológico Interurbano en la microcuenca del río Bermúdez en Heredia, Costa Rica, caracterizando la variación acústica de cada sitio mediante índices de paisaje sonoro que permitieron interpretar la interacción acústica de las biofonías, geofonías y antropofonías a lo largo de la microcuenca con los que se logró evidenciar la relación de las variables acústicas con las variables biológicas. En ese sentido, las vocalizaciones podrían sugerirse como un elemento para establecer semejanzas y dife-rencias entre especies o familias, concordonado con Fernández et al. (2022), para quienes los sonidos que producen los animales, son una herramienta que ha sido ampliamente usada en el estudio de las aves por su papel en la delimitación e identificación de especies; es decir representan un carácter diagnóstico que facilita la discrimi-nación de entidades taxonómicas.
CONCLUSIONES
A partir de las características cuantitativas de las vocalizaciones después de aplicar el algoritmo de clúster jerárquico, las especies que guardaron similitud fueron: Falco peregrinus y F. columbarius, F. rufigularis y F. femoralis; mientras que F. sparverius tuvo más similaridad con las 2 primeras especies. Todas estas especies se diferencian de las especies de la familia Strigidae teniendo aquí Glaucidium brasilianum, G. jardini y G. peruanum más similaridad; asimismo Glaucidium hardyi y Aegolius harrisii guardan más concordancia entre ellos y finalmente Megascops roboratus presenta similaridad con todas las especies del género Glaucidium (coeficiente de aglomeración = 0,67).
En este estudio se analizaron 53 audios por especie, pero quizás en futuras investigaciones se podrían aumentar el número de audios analizados; pero teniendo en cuenta cantos colectados de museos de historia natural ya que sería una forma de asegurarnos de tener cantos estándar para un mejor análisis y además considerar separar las vocalizaciones de machos y hembras.

