










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. vol.26 no.3 Lima ago. 2019
http://dx.doi.org/10.15381/rpb.v26i3.16776
TRABAJOS ORIGINALES
Diversidad y actividad horaria de mamíferos medianos y grandes registrados con cámaras trampa en el Parque Nacional Tingo María, Huánuco, Perú
Diversity and diel activity of medium and large mammals recorded with camera traps in Tingo María National Park, Huánuco, Peru
E. Daniel Cossios*1 ORCID iD: 0000-0003-4188-7632 , Alex Ricra Zevallos2
1 Biosfera Consultores Ambientales, Lima, Perú.
2 Parque Nacional Tingo María – Servicio Nacional de Áreas Naturales Protegidas por el Estado (SERNANP), Tingo María, Perú.
Resumen
La mastofauna del Parque Nacional Tingo María (PNTM) es conocida principalmente por entrevistas a pobladores locales. Este artículo presenta una lista de mamíferos medianos y grandes en base a evaluaciones con cámaras trampa y su verificación con entrevistas. Las evaluaciones se realizaron entre diciembre del 2014 y noviembre del 2018, con un esfuerzo de 2970 trampas cámara/día, consiguiéndose 1640 registros independientes. Se registró 19 especies silvestres, cuatro de las cuales son nuevos registros para el PNTM (la zarigüeyita acuática Chironectes minimus, el margay Leopardus wiedii, la tigrina Leopardus tigrinus y el osito cangrejero Procyon cancrivorus), tres son nuevos para las yungas peruanas (L. wiedii, el ronsoco Hydrochoerus hydrochaeris y la nutria Lontra longicaudis) y dos (H. hydrochaeris y L. wiedii) tienen el récord de altitud en el Perú. Se elaboró patrones de actividad horaria para las nueve especies con más de 10 registros. Ocho de ellos coinciden con lo publicado anteriormente, mientras que el patrón conseguido para el machetero Dynomis branicki es el primero elaborado en condiciones de silvestría. Tanto la presencia de varias especies de predadores como la normalidad de los patrones de actividad encontrados sugieren un buen estado de conservación del bosque en el PNTM. En conclusión, el muestreo con cámaras trampapermitió la obtención de una lista confiable y bastante completa de los mamíferos medianos y grandes de hábitos terrestres del PNTM, además de datos sobre su distribución y ecología.
Palabras clave: Dinomys branicki; inventario; mamíferos terrestres; patrón de actividad; yungas.
Abstract
The mammal fauna of the Parque Nacional Tingo María (PNTM) mainly is known for local people interviews. In this work, using camera traps and its verification by interviews to local people we report medium and large mammals list of the PNTM. Evaluations were realized between December 2014 and November 2018. Sampling effort was of 2970 camera traps/day, with 1640 independent records. Nineteen wild species were recorded, four of which were new reports to the PNTM (the water opossum Chironectes minimus, the margay Leopardus wiedii, the Northern tiger cat Leopardus tigrinus and the crab-eating raccoon Procyon cancrivorus), three were new reports for the Peruvian yungas (L. wiedii, the capybara Hydrochoerus hydrochaeris and the Neotropical otter Lontra longicaudis) and two (H. hydrochaeris and L. wiedii) have been registered as its higher altitude record in Peru. Activity patterns were elaborated for the nine species with more than 10 records. Eight of them are coincided with the previously published. Activity pattern of pacarana (Dynomis branicki) is the first one elaborated in wild conditions. The presence of several predator species and normal activity patterns, would be suggest a good state of forest conservation in the PNTM. In conclusion, the sampling using camera traps allow to obtain a reliable and fairly complete list of the medium and large terrestrial mammals of the PNTM, as well as data on their distribution and ecology.
Keywords: activity pattern; Dinomys branicki; species inventory; terrestrial mammals; yungas.
Introducción
El Parque Nacional Tingo María (PNTM) es la segunda área natural protegida creada en el Perú. Se encuentra a 6.5 km de la ciudad de Tingo María, en la ecorregión de Yungas, la cual se caracteriza por su alta biodiversidad y por sus bosques de altura, húmedos y siempreverdes, situados entre los 800 y 3600 metros de altitud (INRENA 2002). A pesar de su cercanía a una ciudad y de ser de fácil acceso, poca investigación se ha realizado sobre su biodiversidad. En particular, la información publicada sobre la mastofauna ha sido conseguida a partir de entrevistas a pobladores locales (Dourojeanni & Tovar 1972, Gonzales & Llerena 2014), y no de registros obtenidos con métodos directos constituyéndose en un vacío importante.
El uso de cámaras trampa constituye un método directo que permite la observación de especies elusivas o raras con un esfuerzo relativamente pequeño (Rowcliffe et al. 2008), además de permitir la descripción del uso que éstas hacen del hábitat (Head et al. 2012), de sus patrones de actividad (Michalski & Norris 2011, Rowcliffe et al. 2014) y de su abundancia relativa (Jenks et al. 2011), entre otros aspectos de la ecología de las especies registradas.
Aquí, presentamos los resultados de dos evaluaciones hechas con cámaras trampa en el PNTM entre los años 2014 y 2018. Estas evaluaciones tuvieron como objetivo principal el obtener una lista de especies de mamíferos mayores y medianos de hábitos terrestres, que puede servir de base para futuros estudios y para el monitoreo de diversidad biológica en el PNTM, considerándose como especies medianas o grandes aquellas cuyos adultos pesan más de 1 kg (e.g. Botelho et al. 2012, dos Santos & Rodrigues 2013). Adicionalmente, los datos permitieron describir sus patrones de actividad horaria, mejorando nuestro conocimiento sobre la ecología de los mamíferos del PNTM.
Material y métodos
El Parque Nacional Tingo María se ubica en los distritos de Mariano Dámaso Beraún y Rupa Rupa, provincia de Leoncio Prado, departamento de Huánuco, Perú, y se encuentra bordeado por el río Huallaga por el Este y por el río Monzón por el Norte (Fig. 1). Tiene una superficie de 4777 ha y se sitúa entre los 650 y los 1808 m de altitud (INRENA 2002).
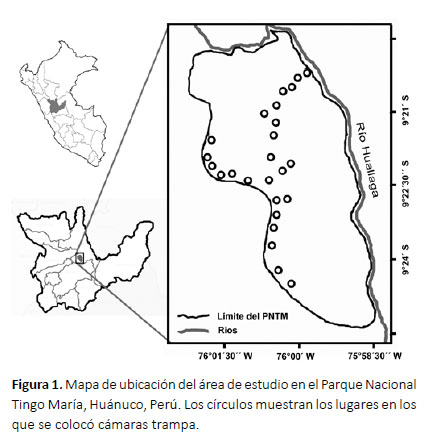
La fase de campo fue realizada en dos periodos: el primero tuvo lugar entre el 8 de noviembre y el 1 de diciembre del 2014, mientras el segundo fue entre el 1 de septiembre del 2017 y el 30 de noviembre del 2018. Durante ambos periodos, entre seis y diez cámaras trampa (Bushnell HD) estuvieron funcionando al mismo tiempo. Ese número varió debido a que no todas las cámaras se encontraron funcionales o disponibles en el mismo momento. Las cámaras fueron cambiadas de sitio cada dos meses con el objetivo de cubrir un mayor número de puntos frecuentados por distintos animales, pero siempre a lo largo de los mismos caminos, principalmente en las partes altas del PNTM. Los puntos en los que las cámaras fueron colocadas pueden apreciarse en la Figura 1.
Las cámaras fueron colocadas a 30 – 40 cm del suelo, en caminos de animales, en zonas despejadas dentro del bosque, apuntando hacia comederos de barro conocidos como collpas o hacia orillas de cursos de agua en lugares que podrían ser usados como cruces por mamíferos. El rango de altitud al que se les colocó fue desde los 680 hasta los 1275 m. Las cámaras fueron programadas para que tomaran tres fotos y un video de 15 segundos por evento, con un periodo de pausa de cinco minutos entre cada evento registrado, y registrando la fecha y hora en cada fotografía y video.
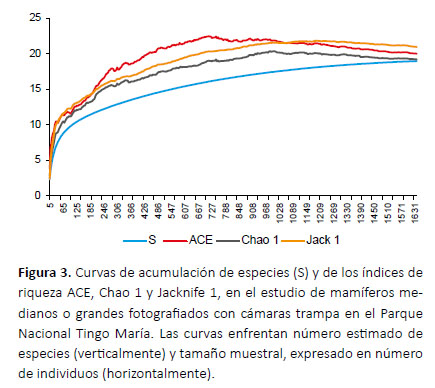
La riqueza real de especies fue estimada con los índices de riqueza ACE, Chao1 y Jacknife1, que utilizan los datos de abundancia y de especies raras en la muestra (Gotelli & Colwell 2001), con el uso del programa EstimateS 9.1.0 (Colwell 2013). Para estimar la confiabilidad de los resultados, se graficó una curva de acumulación de especies por rarefacción y curvas del comportamiento de los índices de riqueza al variar el tamaño de la muestra (Colwell et al. 2012), utilizando el programa Excel y los resultados arrojados por Estimates. Un comportamiento asintótico de las curvas es indicativo de la confiabilidad de los resultados (Gotelli & Colwell 2001).
Para cada especie, se calculó el esfuerzo necesario para registrar el primer evento y la frecuencia de captura. El esfuerzo realizado para lograr el primer registro de cada especie fue expresado en cámaras trampa/día (TCD = la suma de cámaras trampa en funcionamiento por cada día). La frecuencia de captura fue calculada como el número de registros independientes por cada 1000 TCD (Tobler et al. 2008). Se consideró registros independientes a los siguientes casos: a) registros consecutivos de diferentes especies, b) registros consecutivos de la misma especie, tomados en diferentes periodos de una hora y c) registros de la misma especie pero no consecutivos, es decir separados por un registro de otra especie. Debido a que no pudo diferenciarse individuos dentro de cada especie, todas las fotografías de una misma especie tomadas por la misma cámara en el mismo periodo de una hora fueron consideradas como un único registro (Norris et al. 2010).
Cada registro fue clasificado como diurno, nocturno o crepuscular, según el momento en el que fue realizado. El crepúsculo fue definido para cada día como el periodo entre el amanecer astronómico y la hora de salida del sol (crepúsculo matutino o alba), y entre la hora de puesta del sol y el atardecer astronómico (crepúsculo de la tarde), utilizando el programa Moonrise 3.5 (Sidell 2002).
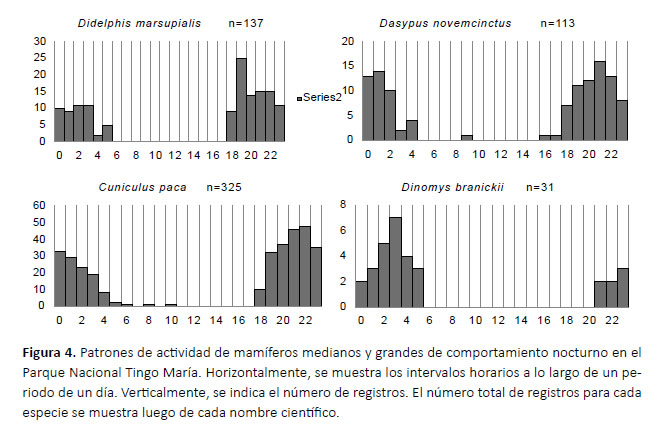
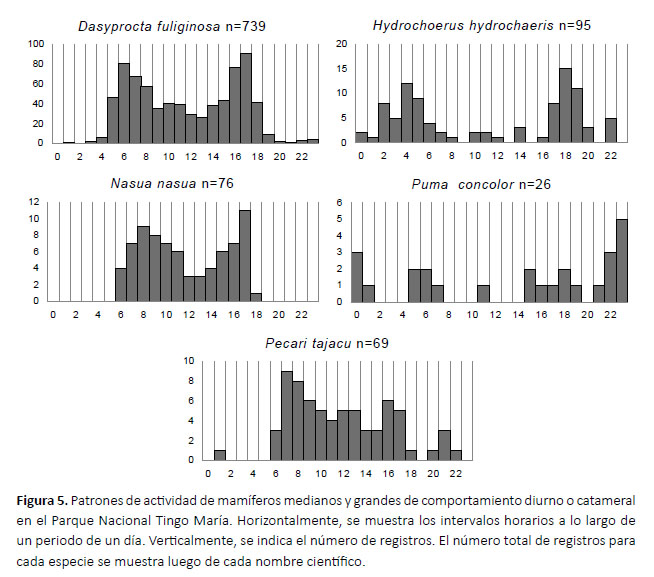
Con el número de registros independientes obtenidos por cada periodo de una hora, se graficó el horario de actividad de cada especie (Rowcliffe et al. 2014) utilizando el programa Excel. Este procedimiento fue realizado únicamente con las especies para las cuales se obtuvo más de 10 registros independientes, por considerarse que un número menor de registros es insuficiente para caracterizar el horario de actividad de una especie (Leuchtenberger et al. 2018).
Resultados
Un esfuerzo de muestreo de 2970 cámaras trampa/ día (TCD) fue llevado a cabo, obteniéndose el registro de 1640 eventos independientes de mamíferos medianos o grandes. Un total de 19 especies de mamíferos silvestres fueron registradas (Tabla 1 y Fig.2), incluyendo dos marsupiales del Orden Didelphimorphia: zarigüeya orejinegra Didelphis marsupialis Linnaeus, 1758 y zarigüeyita acuática Chironectes minimus (Zimmerman, 1780); un Cingulata: armadillo de nueve bandas Dasypus novemcinctus Linnaeus, 1758; un Pilosa: tamandúa Tamandua tetradactyla (Linnaeus, 1758); un Primate: machín blanco Cebus yuracus Hershkovitz, 1949; cinco Rodentia: puerco espín Coendou bicolor (Tschudi, 1844), machetero Dinomys branickii Peters, 1873, ronsoco Hydrochoerus hydrochaeris (Linnaeus, 1766), añuje Dasyprocta fuliginosa Wagler, 1832, y majás Cuniculus paca (Linnaeus, 1766); ocho Carnivora: tigrina Leopardus tigrinus (Schreber, 1775), margay Leopardus wiedii (Schinz, 1821), puma Puma concolor (Linnaeus, 1771), yahuarundí Herpailurus yagouaroundi (É. Geoffroy SaintHilaire, 1803), nutria Lontra longicaudis (Olfers, 1818), manco Eira barbara (Linnaeus, 1758), coatí de cola anillada Nasua nasua (Linnaeus, 1766), y osito cangrejero Procyon cancrivorus; y un Cetartiodactyla: sajino Pecari tajacu (Linnaeus, 1758). Fotografías de roedores y marsupiales pequeños también fueron obtenidas pero no fueron tomadas en cuenta para el presente estudio por no poder identificar su género o especie. Asimismo, se registró al perro doméstico Canis lupus familiaris, en ocho ocasiones.
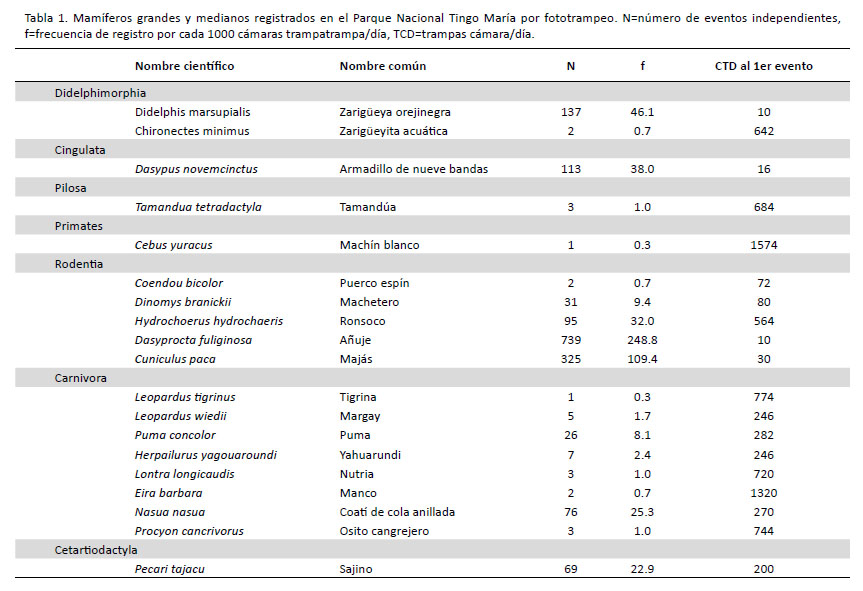

Los índices ACE, Chao1 y Jacknife1 estimaron la riqueza de especies en 20.04, 19.25 y 20.99, por lo que las 19 especies observadas equivaldrían a 90.5 – 98.7% de la riqueza real estimada. La curva de acumulación de especies y las curvas de los índices de riqueza mostraron un comportamiento asintótico (Fig. 3).
El esfuerzo necesario para registrar el primer evento varió entre 10 TCD para la zarigüeya común y 1574 TCD para el caso del machín blanco. La frecuencia de captura más elevada correspondió al añuje, con 248.8 registros/1000 TCD (con 739 registros independientes), mientras que la más baja fue de 0.3 registros/1000 TCD para la tigrina y el machín blanco, que fueron registrados una sola vez cada uno (Tabla 1).
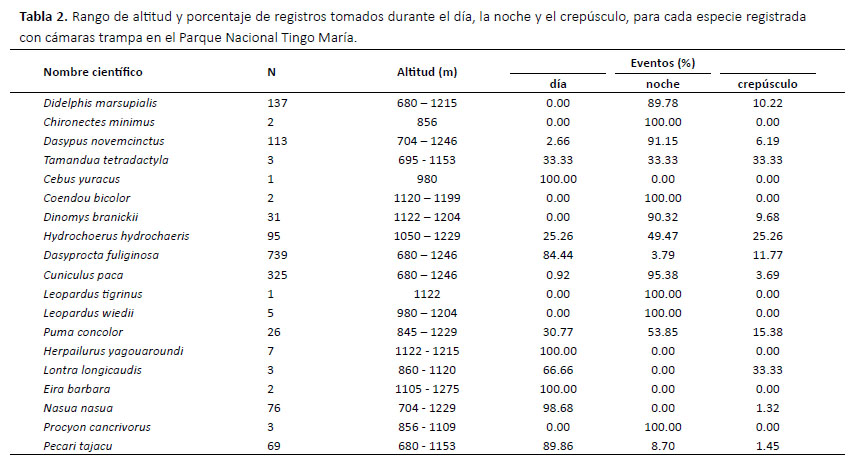
Se consiguió más de 10 registros y se graficó el horario de actividad para nueve de las 19 especies registradas. Cuatro especies (C. paca, D. novemcinctus, D. marsupialis y D. branickii) mostraron actividad principalmente nocturna, con cierta actividad crepuscular y, en los casos del C. paca y D. novemcinctus, pocos registros diurnos (Fig. 4). Tres especies (D. fuliginosa, N. nasua y P. tajacu) fueron principalmente diurnos. H. hydrochaeris y P. con- color mostraron una actividad catemeral (i.e. con cantidades importantes de actividad tanto en la fase luminosa como en la oscura del ciclo diario; Tattersall 1987, 2006), con un pico entre las 23:00 y las 24:00 hrs para P. concolor y picos entre las 04:00-05:00 y 18:00-19:00 hrs para H. hydrochaeris (Fig. 5). El porcentaje de registros conseguidos para cada especie de noche, de dÃa y durante el crepúsculo, asà como el rango de altitud a la que se consiguió los registros, se muestran en la Tabla 2.
Discusión
De las 19 especies de mamíferos medianos y grandes registradas en esta investigación, tres (C. yuracus, T. tetradactyla y C. bicolor) tienen hábitos arborícolas, siendo raramente vistas en el suelo. Su registro en este estudio, con cámaras trampa colocadas cerca al suelo, puede ser tomado como incidental. La baja frecuencia de registro conseguida para estas especies, por lo tanto, no debe ser tomada como indicadora de una baja abundancia de las mismas en el PNTM, sino como una baja probabilidad de registro a nivel del suelo.
Tres publicaciones anteriores presentan listas de mamíferos para el PNTM: Dourojeanni y Tovar (1972) ofrecen una lista hecha mediante entrevistas (salvo una observación directa de ocelote Leopardus pardalis), con 19 especies medianas o grandes y de comportamiento terrestre. INRENA (2002) reporta 17 especies, aunque no informa si sus registros se sustentan en avistamientos directos, entrevistas o suposiciones. La lista de Gonzales y Llerena (2014), también elaborada a partir de entrevistas, reportó la cacería de 16 especies de mamíferos medianos y grandes en las zonas de uso especial y de amortiguamiento del PNTM, siendo algunas de dichas especies registradas solo a nivel de género (Leopardus sp., Cebus sp., Saimiri sp.).
Considerando esas publicaciones, nuestra evaluación con cámaras trampa aporta el registro de cuatro especies no antes reportadas en el PNTM: C. minimus, L. wiedii, L. tigrinus y P. cancrivorus. Además, las cámaras permiten verificar la altitud de los registros y dan certeza a los mismos, permitiendo la identificación a nivel de especie y evitando identificaciones dudosas. Los indicadores de riqueza muestran que las 19 especies observadas se acercan bastante al total de especies esperadas en el PNTM, lo que sugiere que el muestreo fue bastante completo.
En el Perú, L. wiedii ha sido anteriormente registrado entre los 210 y los 860 m (MINAM 2011), mientras que H. hydrochaeris ha sido registrado hasta los 900 m (Pacheco et al. 2018). Los registros aquí reportados para estas dos especies, entre 980 y 1204 m para L. wiedii y entre 1050 y 1229 m para H. hydrochaeris, son entonces los de mayor altitud en el país actualmente. Los registros de H. hydrochaeris, L. wiedii y L. longicaudis son también notables por ser los primeros para las yungas peruanas (MINAM 2011, Pacheco et al. 2009).
La modificación de los patrones de actividad horaria puede ser indicadora del impacto de perturbaciones antropogénicas. Así, la caza y la actividad turística pueden perturbar el horario de actividad de ciertas especies, disminuyendo su actividad durante las horas en las que la presencia humana es mayor, generalmente de día, y acrecentándola cuando es menor (e.g. Keuling et al. 2008, Little et al. 2016, Marchand et al. 2014). Los patrones de actividad horaria aquí descritos de D. marsupialis, D. novemcinctus, D. fuliginosa, C. paca, H. hydrochaeris, N. nasua y P. tajacu concuerdan con observaciones en otras localidades de su distribución (Albanesi et al. 2016, Ávila-Nájera et al. 2016, Huck et al. 2017, Leuchtenberger et al. 2018, Mosquera-Guerra et al. 2018, Parodi 2015). Aunque existe actividad cinegética (Gonzales & Llerena 2014) y turística (INRENA 2002) dentro del PNTM y su zona de amortiguamiento, nuestros resultados sugieren que el comportamiento de dichas especies no está siendo afectado de forma notable por la actividad humana en el parque, en lo que se refiere a la actividad horaria.
La deforestación es otro factor que puede afectar el patrón de actividad horaria de una especie, como ha sido reportado para D. novemcinctus, que aumenta drásticamente su actividad diurna en ambientes con bosques distribuidos en parches (Norris et al. 2010). El comportamiento registrado aquí para el armadillo sería un indicador del buen estado de conservación del bosque en el PNTM.
En el caso de P. concolor, aunque en otras publicaciones se muestra, como aquí, su comportamiento catemeral, los picos de actividad de esta especie difieren en distintos lugares estudiados. Mientras nuestros resultados muestran un pico de actividad entre las 22:00 y la 01:00 horas, otros autores los muestran a las 04:00, 16:00, 17:00, 18:00 o 24:00 horas (Foster et al. 2013, Maffei et al. 2002, Parodi 2015). Varios estudios indican que el patrón de actividad de P. concolor está determinado por la actividad de sus presas (Emmons 1987, Harmsen et al. 2000, Nuñez et al. 2000), por lo que esta variación en comportamiento es considerada normal.
La actividad horaria de D. branicki ha sido estudiada antes en condiciones de cautiverio, mostrando picos de actividad a las 18:00, 23:00 y 01:00 horas (López et al. 2000). Hasta donde sabemos, nuestros resultados son los primeros sobre la actividad horaria de esta especie en condiciones naturales, y difieren del estudio en cautiverio al mostrar un solo pico, a las 03:00 h, y ningún registro antes de las 21:00 h (Fig. 4). Esta diferencia podría deberse a múltiples factores, como distintas condiciones ambientales (i.e. iluminación, temperatura, lluvia, etc.), presencia de predadores y especies competidoras.
Tanto la presencia de varias especies de predadores como la normalidad en los patrones de actividad observadas en las especies estudiadas sugieren un buen estado de conservación del bosque en el PNTM. Sin embargo, la falta de registros de algunas especies de mamíferos grandes como el venado cola blanca Odocoileus peruvianus, el venado rojo Mazama americana, el jaguar Panthera onca, el tapir Tapirus terrestris y el ocelote Leopardus pardalis, que sí fueron registradas, por entrevistas, por Dourojeanni y Tovar (1972) y cuya presencia en la zona es esperable debido al tipo de hábitat existente, puede ser indicador de una disminución de sus poblaciones. Dado que se espera muy pocas adiciones a nuestra lista de mamíferos terrestres medianos o grandes, suponemos que estas especies, si es que ocurren en el PNTM, tengan una presencia ocasional en la actualidad. El área del PNTM es relativamente pequeña y es probablemente insuficiente para asegurar el mantenimiento de poblaciones de dichas especies, por lo que el mantenimiento de buenas condiciones de conservación en las áreas aledañas sería fundamental para eso.
En conclusión, el muestreo con cámaras trampa permitió la obtención de una lista confiable y bastante completa de los mamíferos medianos y grandes de hábitos terrestres dentro del PNTM, además de nuevos datos sobre su distribución y ecología. Es recomendable realizar evaluaciones periódicas con este método para verificar si la presencia de especies raras continúa en el futuro y si otras especies de mamíferos grandes se presentan de forma ocasional.
Agradecimientos:
Agradecemos muy especialmente a los guardaparques Edilberto Eugenio Bravo, Henry Tolentino Diego, Hernán Cornelio Adán, Luis Adriano Espinoza, Cesar Rosas Aparicio y Emiliano Carrillo Mena, y a Bessy Cobos Panduro, especialista del PNTM, que ayudaron en la instalación y revisión de cámaras, proporcionando datos valiosos y consejos sobre los lugares a visitar, y a Tupac, Rumi y Chaska Cossios, que participaron en la ubicación inicial de sitios de muestreo.
Rol de los autores:
EDC organizó el muestreo, participó en el trabajo de campo y realizó el análisis de datos. ARZ organizó y participó en el trabajo de campo.
Ambos autores revisaron y aprobaron el manuscrito.
Conflicto de intereses:
Los autores no incurren en conflictos de intereses.
Fuentes de financiamiento:
La evaluación del 2014 se realizó gracias al financiamiento de Empresa de Generación Huallaga SA.
La evaluación del 2017-2018 contó con financiamiento propio y de Biosfera Consultores Ambientales SAC.
Aspectos éticos / legales:
Permiso de investigación N°RJ-PNTM-007-2017-SERNANP-JEF
Citación:
Cossios E.D. y A. Ricra Zevallos. 2019. Diversidad y actividad horaria de mamíferos medianos y grandes registrados con cámaras trampa en el Parque Nacional Tingo María, Huánuco, Perú. Revista peruana de biología 26(3): 325 332 (Septiembre 2019). doi: http://dx.doi.org/10.15381/rpb.v26i3.16776
Literatura citada
Albanesi S.A., J.P. Jayet & A.D. Brown. 2016. Patrones de actividad de mamíferos de medio y gran porte en el pedemonte de yungas del noroeste argentino. Mastozoología Neotropical. 23(2):335-358. [ Links ]
Ávila-Nájera D.M., C. Chávez, M.A. Lazcano-Barrero, G.D. Mendoza & S. Perez-Elizalde. 2016. Overlap in activity patterns between big cats and their prey in northern Quintana Roo, Mexico. Therya. 7(3):439-448. doi: https://doi.org/10.12933/therya-16-379 ISSN 2007-3364. [ Links ]
Botelho A.L.M., A.M. Calouro, L.H.M. Borges & W.Z. Chaves. 2012. Large and medium-sized mammals of the Humaitá Forest Reserve, southwestern Amazonia, state of Acre, Brazil. Check List. 8(6):1190-1195. doi: https://doi.org/10.15560/8.6.1190. [ Links ]
Colwell R.K. 2013. EstimateS, Version 9.1: Statistical Estimation of Species Richness and Shared Species from Samples (Software and User's Guide). Freeware for Windows and Mac OS. [ Links ]
Colwell R.K., A. Chao, N.J. Gotelli, S.Y. Lin, C.X. Mao, R.L. Chazdon & J.T. Longino. 2012. Models and estimators linking individual-based and sample-based rarefaction, extrapolation, and comparison of assemblages. Journal of Plant Ecology. 5:3-21. doi: https://doi.org/10.1093/jpe/rtr044. [ Links ]
dos Santos P.V., F. Rodrigues. 2013. Assemblage of medium and large size mammals in an urban semideciduous seasonal forest fragment in cerrado biome. Biota Neotropical. 13(2):76-80. doi: https://doi.org/10.1590/S1676-06032013000200008 [ Links ]
Doujojeanni M, Tovar A. 1972a. Evaluación y bases para el manejo del Parque Nacional de Tingo María (Huánuco, Perú). Informe Especial. Universidad Nacional Agraria. 71 pp [ Links ]
Emmons L.H. 1987. Comparative feeding ecology of felids in a neotropical rainforest. Behaviour Ecology and Sociobiology. 20:271-283. doi: https://doi.org/10.1007/BF00292180. [ Links ]
Foster V.C., P. Sarmento, R. Sollman, N. Torres, A.T. Jácomo, N. Negroes, C. Fonseca & L. Silveira. 2013. Jaguar and puma activity patterns and predator-prey interactions in four Brazilian biomes. Biotropica. 45(3):373-379. doi: https://doi.org/10.1111/btp.12021. [ Links ]
Gonzales F.N. & G. Llerena. 2014. Cacería de mamíferos en la zona de uso especial y de amortiguamiento del Parque Nacional Tingo María, Huánuco, Perú. Revista Peruana de Biología. 21(3):283-286. doi: https://doi.org/10.15381/rpb.v21i3.10904. [ Links ]
Gotelli N. & R.K. Colwell. 2001. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters. 4: 379-391. doi: https://doi.org/10.1046/j.1461-0248.2001.00230.x [ Links ]
Harmsen B.J., J. Foster, S.C. Silver, L.E.T. Ostro & C.P. Doncaster. 2011. Jaguar and puma activity patterns in relation to their main prey. Mammalian Biology. 76:320-324. doi: https://doi.org/10.1111/btp.12021. [ Links ]
Head J.S., M.M. Robbins, R. Mundry, L. Makaga & C. Boesch. 2012. Remote video-camera traps measure habitat use and competitive exclusion among sympatric chimpanzee, gorilla and elephant in Loango National Park, Gabon. Journal of Tropical Ecology. 28:571-583. doi: https://doi.org/10.1017/50266467412000612. [ Links ]
Huck M., C.P. Juarez, M.A. Rotundo, V.M. Dávalos & E. FernandezDuque. 2017. Mammals and their activity patterns in a forest area in the humid chaco, Northern Argentina. Check List. 13(4):363-378. doi: https://doi.org/10.15560/13.4.363. [ Links ]
INRENA (Instituto Nacional de Recursos Naturales). 2002. Plan maestro 2003 – 2007, Parque Nacional Tingo María. Tingo María-Perú. 90p. [ Links ]
Jenks K.E., P. Chanteap, K. Damrongchainarong, P. Cutter, P. Cutter, T. Redford, A.J. Lynam, J. Howard & P. Leimgruber. 2011. Using relative abundance indices from cameratrapping to test wildlife conservation hypotheses – an example from Khao Yai National Park, Thailand. Tropical Conservation Science. 4(2):113-131. doi: https://doi.org/10.1177/194008291100400203. [ Links ]
Keuling O., N. Stier & M. Roth. 2008. How does hunting influence activity and spatial usage in wild boar Sus scrofa L.? European Journal of Wildlife Research. 54:729-737. doi: https://doi.org/10.1007/s10344-008-0204-9. [ Links ]
Leuchtenberger C., E.S. de Oliveira, L.P. Cariolatto & C.B. Kasper. 2018. Activity patterns of medium and large sized mammals and density estimates of Cuniculus paca (Rodentia: Cuniculidae) in the Brazilian pampa. Brazilian Journal of Biology. 78(4):697-705. doi: https://doi.org/10.1590/1519-6984.174403. [ Links ]
Little A.R., S.L. Webb, S. Demarais, K.L. Gee, S.K. Riffell & J.A. Gaskamp. 2016. Hunting intensity alters movement behavior of white-tailed deer. Basic and Applied Ecology. 17:360-369. doi: https://doi.org/10.1016/j.baae.2015.12.003. [ Links ]
López L., I. López, J. Mora & K. Osbahr. 2000. Estudio preliminar del comportamiento de Dinomys branickii (Peters, 1873) en cautiverio. Revista. U.D.C.A Actividad & Divulgación Científica. 3(1):28-35. [ Links ]
Maffei L., E. Cuellar & A.J. Noss. 2002. Uso de trampas-cámara para la evaluación de mamíferos en el ecotono chacochiquitanía. Revista Boliviana de Ecología. 11:55-65. [ Links ]
Marchand P., M. Garel, G. Bourgoin, D. Dubray, D. Maillard & Loison. 2014. Impacts of tourism and hunting on a large herbivore´s spatio-temporal behavior in and around a French protected area. Biological Conservation. 177:1-11. doi: https://doi.org/10.1016/j.biocon.2014.05.022.
Michalski F. & D. Norris. 2011. Activity pattern of Cuniculus paca (Rodentia: Cuniculidae) in relation to lunar illumination and other abiotic variables in the southern Brazilian Amazon. Zoologia. 28(6):701-708. doi: https://doi.org/10.1590/S1984-46702011000600002. [ Links ]
MINAM (Ministerio del Ambiente del Perú). 2011. Estudio de especies CITES de carnívoros peruanos. Informe final. 162 pp. [ Links ]
Mosquera-Guerra F., F. Trujillo, A.P. Diaz-Pulido & H. MantillaMeluk. 2018. Diversidad, abundancia relativa y patrones de actividad de los mamíferos medianos y grandes asociados a los bosques riparios del río Bita, Vichada, Colombia. Biota Colombiana. 19(1):202-218. doi: https://doi.org/10.21068/c2018v19n01a13an01. [ Links ]
Norris D., F. Michalski & C.A. Peres. 2010. Habitat patch size modulates terrestrial mammal activity patterns in Amazonian forest fragments. Journal of Mammalogy. 91:551-560. doi: https://doi.org/10.1644/09MAMM-A-199.1. [ Links ]
Nuñez R., B. Miller B & F. Lindzey. 2000. Food habits of jaguars and pumas in Jalisco, Mexico. Journal of Zoology. 252:373379. doi: https://doi.org/10.1111/j.1469-7998.2000.tb00632.x. [ Links ]
Pacheco V., B. Inche & W. Wust. 2018. Mamíferos del Perú. Grupo La República Publicaciones S.A. 120 pp. [ Links ]
Pacheco V., R. Cadenillas, E. Salas, C. Tello & H. Zeballos. 2009. Diversidad y endemismo de los mamíferos del Perú. Revista Peruana de Biología 16(1):5-32. doi: https://doi.org/10.15381/rpb.v16i1.111. [ Links ]
Parodi A. 2015. Patrones de actividad e influencia del ciclo lunar en la actividad de una comunidad animal del Parque Nacional del Manu. Tesis para optar el título de Licenciado en Biología. Universidad Peruana Cayetano Heredia. [ Links ]
PayanE.&T.deOliveira.2016.Leopardustigrinus.TheIUCNRedList of Threatened Species 2016: http://dx.doi.org/10.2305/IUCN.UK.2016-2.RLTS.T54012637A50653881.en. Downloaded on 18 March 2019. [ Links ]
Rowcliffe J.M., J. Field, S.T. Turvey & C. Carbone. 2008. Estimating animal density using camera traps without the need for individual recognition. Journal of Applied Ecology. 45:1228-1236. doi: https://doi.org/10.1111/j.13652664.2008.01473.x. [ Links ]
Rowcliffe J.M., R. Kays, B. Kranstauber, C. Carbone & P.A. Jansen. 2014. Quantifying levels of animal activity using camera trap data. Methods in Ecology and Evolution. 201(5):1170-1179. doi: https://doi.org/10.1111/2041-210X.12278. [ Links ]
Sidell B.P. 2002. Moonrise 3.5 (32 bits). Software. Tattersall I. 1987. Cathemeral activity in primates: a definition. Folia Primatologica. 49:200-202. doi: https://doi.org/10.1159/000156323. [ Links ]
Tattersall I. 2006. The concept of cathemerality: history and definition. Folia primatologica. 77:7-14. doi: https://doi.org/10.1159/000089692. [ Links ]
Tobler M.W., S.E. Carrillo-Percastegui & G. Powell. 2009. Habitat use, activity patterns and use of mineral licks by five species of ungulate in south-eastern Peru. Journal of Tropical Ecology, 25(3): 261-270. doi: https://doi.org/10.1017/S0266467409005896. [ Links ]
Presentado: 08/04/2019
Aceptado:26/07/2019
Publicado online:30/09/2019

