











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
La infiltración es el movimiento del agua de la atmósfera al suelo a través de una interfaz definible pero intangible (Surytmojo, 2014). La infiltración del agua en el suelo es un proceso importante del ciclo del agua en los ecosistemas terrestres, debido a que este influye en el balance hídrico de la vegetación, la perdida potencial de la capa superior del suelo por erosión, la escorrentía y la recarga de las aguas subterráneas (Sun et al., 2018; Ren et al., 2021). Además, la capacidad de infiltración del suelo está influida por las características del suelo, las estructuras, los factores químicos y los factores biológicos (Luo et al., 2014; Wang et al., 2018; Tapia et al., 2020; Bork et al., 2021).
Por otro lado, la capacidad de infiltración puede ser modificada de forma rápida por las actividades antrópicas (López et al., 2018). La rápida expansión urbana y agrícola ocasionan impactos severos en la capacidad de retención de agua en los suelos (Galli et al., 2021; Santos et al., 2021), además la actividad ganadera y los incendios forestales pueden reducir significativamente la capacidad de infiltración de los suelos (Karlin et al., 2019; Fernández et al., 2021).
Estudios previos han demostrado que la cobertura forestal tiene influencia significativa en la reducción de la erosión de los suelos (Ren et al., 2021), la escorrentía y las inundaciones (Regüés et al., 2017). Por lo que un cambio en el uso del suelo puede alterar el balance del ciclo hidrológico y afectar de forma negativa la infiltración y el escurrimiento superficial (Lozano‑Trejo et al., 2020). Por lo tanto, la infiltración suele ser menor en áreas sin cobertura vegetal (suelo desnudo) y en áreas agrícolas, en comparación con las de cobertura forestal (de Almeida et al., 2018; Tang et al., 2019). Sin embargo, en la Amazonia peruana, a nuestro conocimiento, no se ha investigado en detalle los impactos de los cambios en la cobertura vegetal en la infiltración de los suelos. Por lo que son necesarios este tipo de estudios para comprender las implicancias de estos cambios en los ciclos hidrológicos (Galli et al., 2021).
En la Amazonia peruana, específicamente en Madre de Dios, desde mediados del siglo pasado una gran extensión de bosque ha sido deforestado con fines agrícolas, ganaderos y por la minería de oro, especialmente en zonas cercanas a vías de acceso como la carretera interoceánica (Nicolau et al., 2019; Tarazona & Miyasiro‑López, 2020; Alarcón et al., 2021). La agricultura migratoria es considerada como una de las principales fuentes de deforestación (Alarcón et al., 2021), además como producto de estas actividades antrópicas en la Amazonia de Madre de Dios, se tiene pastizales abandonados que son más de 100 mil ha en los últimos 20 años (Caballero et al., 2018). Estos cambios en la cobertura vegetal pueden estar afectando las propiedades del suelo en diferentes maneras.
El objetivo del presente estudio fue comparar la capacidad y velocidad de infiltración entre los dos tipos de cobertura vegetal (pastizales y bosques primarios) y analizar la relación entre la capacidad de infiltración con la abundancia, riqueza y diversidad florística en la Amazonia peruana (Madre de Dios).
MATERIALES Y MÉTODOS
ÁREA DE ESTUDIO
El lugar de estudio está ubicado en el sector de San Bernardo, en las coordenadas geográficas 12°44'17,18"S ‑ 69°23'52,14"O perteneciente al distrito de Tambopata y Laberinto (Madre de Dios), al suroeste de la Amazonía peruana. La precipitación media anual y la temperatura media son 2120 mm y 25,4 °C (Garate‑Quispe et al., 2020) respectivamente (considerando el periodo 1980‑2016). El sitio de estudio presenta una estación seca (junio a setiembre (Garate‑Quispe et al., 2021) cuando la precipitación es menor a 100 mm.
INFILTRACIÓN
Se realizaron 6 mediciones de infiltración en bosque primario y 3 en pastizal. La tasa de infiltración se determinó utilizando un infiltrómetro de anillo simple (Sepúlveda, 1999), las dimensiones del anillo fueron de 17 cm de diámetro exterior y 20 cm de altura. El infiltrómetro se introdujo 2 cm de profundidad y se lo llenó con agua hasta un nivel de 20 cm por encima de la superficie del suelo (Figura 1). Para calcular la tasa de infiltración y la infiltración acumulada se registraron los cambios en el nivel del agua en incrementos de tiempo, hasta que la tasa de infiltración se estabilice utilizando la siguiente ecuación:

Donde:
ΔH = Disminución en el nivel del agua en el infiltrómetro (mm) en un intervalo de tiempo determinado.
t = Tiempo de infiltración del agua en el suelo (minuto)
Los anillos se recargaron después de cada registro.

Figura 1 Diagrama del infiltrómetro de anillo simple utilizado en el estudio. (A) Dimensiones del infiltrómetro. (B) Fotografía de un infiltrómetro en campo (Vagen et al., 2010)
VELOCIDAD DE INFILTRACIÓN
Para el cálculo de la velocidad de infiltración se tomó en cuenta la ecuación modificada de Porta et al. (1994):
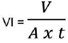
Donde:
VI: Velocidad de infiltración (mm/hora)
V: Volumen de agua en el infiltrómetro (m3)
A: Área interna del infiltrómetro (m2)
t = Tiempo de infiltración (hora)
CARACTERÍSTICAS FISICOQUÍMICAS DEL SUELO
Se colectaron muestras del suelo en dos ti‑ pos de cobertura vegetal: bosque primario intervenido (n=6) y pastizal (n=3). El muestreo se realizó siguiendo las recomendaciones de la Guía de Muestreo de Suelos del Perú (MINAM, 2014). Cada muestra de suelo estuvo compuesta de 6 submuestras superficiales (0‑20 cm) distribuidas de forma sistemática en la cercanía al punto de evaluación de la vegetación. Las muestras de suelo se caracterizaron fisicoquímicamente en el Laboratorio de Suelos y Fertilidad de la Universidad Nacional Agraria La Molina. En la caracterización fisicoquímica se consideró a: porcentaje de materia orgánica (MO), porcentaje de arena, limo, arcilla, concentraciones de fosforo, potasio y capacidad de intercambio catiónico (CIC).
COMPOSICIÓN FLORÍSTICA
Para el muestreo de comunidades de plantas leñosas y pastizales, se utilizó la metodología de Marco de Vigilancia de la Degradación de la Tierra (LDSF) (Vagen et al., 2010), por ser menos propenso a sesgos donde las plantas no están distribuidas aleatoriamente. La metodología LDSF establece un área circular de evaluación de 1000 m2 y dentro de esta área se establece 4 subparcelas circulares de 100 m2 distanciadas entre sı́ por al menos 10 m. En cada subparcela se registraron e identifica‑ ron a todas las plantas leñosas, arbustos (1,5 m‑3 m de altura) y arboles (>3 m de altura).
ANÁLISIS ESTADÍSTICO
Se utilizaron estadísticos descriptivos (promedio ± desviación estándar) para la caracterización fisicoquímica del suelo. Para la comparación de las características fisicoquímicas del suelo e infiltración total promedio según tipo de cobertura, se utilizó la prueba de t‑Student, con un nivel de significancia del 5%. Se construyeron los gráficos de velocidad de infiltración y se ajustó al mejor modelo de regresión, utilizando los datos promedios de la infiltración para ambos tipos de cobertura con el software TableCurve 2D y R. Se analizó la correlación entre la capacidad de infiltración con la abundancia, riqueza y diversidad arbórea utilizando el coeficiente de correlación de Pearson. Para los análisis de correlación se junta‑ ron los datos de los dos tipos de cobertura. Para agrupar los sitios de muestreo en función de la composición florística, se realizó un dendrograma jerárquico (UPGMA), mediante el método de distancias de disimilitud de Jaccard. Para comprobar diferencias significativas entre la estructura jerárquica de los grupos se utilizó la prueba de permutación del perfil de similitud (SIMPROF) usando el software PRIMER 7 (versión 7.0.022). Todas las figuras se realizaron utilizando los paquetes Vegan y ggplot2, disponibles en R (versión 4.2), utilizando el entorno integrado de RStudio (versión 2022.02).
RESULTADOS
CARACTERÍSTICAS FISICOQUÍMICAS DEL SUELO
No se encontraron diferencias significativas en las características fisicoquímicas del suelo entre los dos tipos de cobertura (t‑Student, p > 0,05, Tabla 1). En ambos tipos de cobertura las características texturales fueron similares (Tabla 1), se encontró un predominio del tipo de textura franco y franco‑arenoso, aunque en un solo sitio en pastizal se encontró el tipo franco‑ arcillo‑arenoso. Mientras que la capacidad de intercambio catiónico y los porcentajes de materia orgánica fueron ligeramente superior en el bosque primario que en el pastizal (Tabla 1).
Tabla 1 Comparación de las características fisicoquímicas del suelo entre los dos tipos de cobertura evaluados. MO: Materia orgánica; CIC: Capacidad de Intercambio Catiónico. Prueba de t-Student al 5% de probabilidad
| Tipo de cobertura | MO (%) | Arena (%) | Limo (%) | Arcilla (%) | P (ppm) | K (ppm) | CIC |
|---|---|---|---|---|---|---|---|
| Bosque primario | 2,6 ± 1,3 a | 57,3 ± 14,7 a | 26,2 ± 9,7 a | 16,5 ± 8,3 a | 1,6 ± 1,0 a | 65,0 ± 22,7 a | 5,16 ± 2,9 a |
| Pastizal | 2,1 ± 0,7 a | 56,5 ± 7,4 a | 26,9 ± 11,4 a | 16,5 ± 5,1 a | 1,0 ± 0,8 a | 84,7 ± 31,2 a | 3,81 ± 1,5 a |
COMPOSICIÓN FLORÍSTICA
En total se registraron 52 especies, considerando los 8 puntos de muestreo (bosques y pastizales). En los pastizales encontramos siete especies arbóreas (promedio 3 n± 2 por punto de muestreo), mientras que en los bosques se encontraron 18 (± 3) especies en promedio en cada punto de muestreo (Tabla 2). En pastizales el 71% de las especies corresponden a especies pioneras, de estas las más frecuentes fueron Ochroma pyramidale, Apeiba membranacea y Trema micrantha.
Tabla 2 Abundancia, riqueza y diversidad florística en los sitios evaluados según tipo de cobertura vegetal.
| Abundancia | Riqueza de especies | Diversidad de Shannon | |
|---|---|---|---|
| Bosque 1 | 19 | 19 | 3.42 |
| Bosque 2 | 17 | 17 | 3.30 |
| Bosque 3 | 13 | 13 | 3.03 |
| Bosque 4 | 19 | 19 | 3.42 |
| Bosque 5 | 25 | 21 | 3.38 |
| Pastizal 1 | 3 | 3 | 1.43 |
| Pastizal 2 | 1 | 1 | 0.00 |
| Pastizal 3 | 5 | 5 | 2.01 |
En el bosque primario las especies encontradas más frecuentes (>65% de los puntos evaluados) fueron Oenocarpus mapora, Miconia tomentosa, Eschweilera coriaceae, Guarea macrophylla, Matisia malacocalyx, Nectandra globosa, Pouteria torta, Tachigali paniculata y Virola calophylla. El análisis SIMPROF (Figura 2), mostró la existencia de dos conglomerados o grupos diferenciados de forma significativa por su composición florística (p< 0,05), el primer grupo corresponde a las parcelas ubicadas en el pastizal y el otro grupo corresponde a las parcelas ubicadas en el bosque. Por lo tanto, encontramos una baja similitud florística entre el bosque y el pastizal (Figura 2), teniendo solo dos especies compartidas (Inga capitata y Vismia tomentosa).

Figura 2 Similitud en la composición florística entre el bosque primario y el pastizal, mediante el análisis de conglomerado jerárquico (UPGMA) obtenido a partir de una matriz de similitud de Jaccard. En diferentes colores se muestran los sitios agrupados en 2 conglomerados con significación estadística (SIMPROF).
INFILTRACIÓN
La capacidad de infiltración de agua en el suelo fue significativamente superior en el bosque que en el pastizal (t‑Student, p<0,01) (Figura 3). En promedio, la capacidad de infiltración en los bosques (1162 ± 448 mm/hora) fue 16 veces más que en el pastizal (73 ± 60 mm/hora).

Figura 3 Capacidad de Infiltración total promedio para cada tipo de cobertura vegetal (pastizal y bosque). Los bigotes representan la desviación estándar.
Los valores de capacidad de infiltración estuvieron relacionados de forma directa y significativa con la abundancia (r = 0,83; p ≤ 0,01), riqueza de especies (r = 0,82; p ≤ 0,01) y diversidad de especies (índice de Shanon, r = 0,73; p < 0,01).
Respecto a las curvas de infiltración, encontramos un diferente comportamiento entre los bosques (Figura 4A) y los pastizales (Figura 4B). En los bosques, se observó una mayor velocidad de infiltración en los primeros 10 minutos, seguidamente se observó una disminución en la velocidad de infiltración, entre los 10‑15 minutos, luego esta disminución se vuelve menos acentuada (Figura 4A). Asimismo, a partir de los 35 minutos alcanza la infiltración base, la cual corresponde a la porción asintótica de la curva. En el pastizal, la velocidad de infiltración fue más homogénea y no observamos el descenso rápido que se encontró en los bosques (Figura 4B).

Figura 4 Comportamiento de la infiltración de agua en el suelo según tipo de cobertura vegetal, bosque primario (A) y en pastizales (B).
El modelo de mejor ajuste para la velocidad de infiltración en el bosque fue el modelo potencial (R2 = 0,99, p < 0.05) y en el pastizal, el mejor modelo ajustado fue el inverso del primer orden (R2 = 0,77, p < 0.05) (Figura 5)

DISCUSIÓN
CARACTERÍSTICAS FISICOQUÍMICAS DEL SUELO
Sobre las características fisicoquímicas del suelo en bosques, los niveles de materia orgánica y capacidad de intercambio catiónico (CIC) fueron inferiores a los reportados para Madre de Dios (Velásquez et al., 2021), sin embargo, los porcentajes de arena, limo y arcilla si fueron similares de los reportados por Velásquez et al. (2021). En pastizales, los valores encontrados de materia orgánica (1,41%), porcentaje de arena (48%), capacidad de intercambio catiónico (83 meq/100g) y potasio (61 ppm) fueron similares a los reportados por Soto‑Benavente et al. (2020) en pastizales en Madre de Dios. Sin embargo, Schwartzmann (2019) reportó en pastizales valores promedio de capacidad de intercambio catiónico (CIC = 17,8 meq/100g), materia orgánica (2,69%), porcentajes de limo (52%) y arcilla (34%) superiores al presente estudio. Los altos valores de capacidad de intercambio catiónico encontrado por Schwartzmann (2019) en pastizales de Madre de Dios se debería a los altos valores de arcilla y materia orgánica registrados, ya que el CIC está influenciado por el contenido de materia orgánica (Brasil et al., 2021) y del tipo de arcilla como de su contenido (Meimaroglou & Mouzakis, 2019).
COMPOSICIÓN FLORÍSTICA
Sobre la vegetación, en el bosque, las especies más frecuentes correspondieron a las típicas familias de bosques primarios: Arecaceae, Lecythidaceae, Meliaceae y Sapotaceae. Estas familias son consideradas hiper‑dominantes en la Amazonia (Draper et al., 2021) y en Madre de Dios (Dueñas & Garate, 2018). Asimismo, conforme a lo esperado en el bosque, se encontraron especies típicas de los bosques primarios remanentes como las del género Oenocarpus (palmeras), Eschweiteria, Guarea, Nectandra y Pouteria (de Assis et al., 2017). Por otro lado, en la Amazonia, la colonización inicial de áreas degradadas o abandonadas suele estar dominada por especies arbóreas pioneras, exigentes de luz, de crecimiento rápido en diámetro y área basal (Chazdon et al., 2007; van Breugel et al., 2007, Vieut et al., 2013). Especies como Ochroma pyramidale, Apeiba membranacea y Trema micrantha son típicas en la sucesión secundaria de áreas abandonadas por actividades antrópicas en Madre de Dios (Román‑Dañobeytia et al., 2015; Garate‑Quispe et al., 2021).
INFILTRACIÓN
Lopes et al. (2008) en un estudio en la Amazonia brasileña, encontraron que la infiltración de agua en el suelo es significativamente superior en presencia de vegetación arbórea. En el presente estudio, la capacidad de infiltración en el pastizal fue significativamente inferior a la del bosque primario, similares resultados fue‑ ron reportados por estudios previos (Regüés et al., 2017; Sun et al., 2018; Lozano‑Trejo et al., 2020). Esto debido a que los cambios en el uso del suelo pueden influir en cambios en las propiedades físicas, químicas y biológicas del suelo, y estas determinan la capacidad de infiltración (Barquéz et al., 2014; Lopes et al., 2018; Luna et al., 2020; Tapia et al., 2020). Las actividades agrícolas y ganaderas desarrolladas posterior a la deforestación y abandono del pastizal, pueden interactuar e influir negativa‑ mente en la infiltración del agua y hacerlos más susceptibles a la erosión (Wu et al., 2016; de Almeida et al., 2018).
Las características de la vegetación tiene efectos importantes en la capacidad de infiltración del suelo (Wang et al., 2018). Asimismo, reportaron que la vegetación arbórea puede favorecer la infiltración mediante la ampliación de su zona radicular (Zhang et al., 2021). La fuerte correlación entre la infiltración de agua en el suelo y los atributos florísticos encontrada en el presente estudio nos sugiere que los sitios con mayor abundancia, riqueza y diversidad tienen una mayor capacidad de infiltración en el suelo. Esto se debería a que la vegetación arbórea de los bosques le brindaría una mayor porosidad, permeabilidad y conductividad hidráulica saturada al suelo (Wang et al., 2009; Luo et al., 2014; Galli et al., 2021) a diferencia de los pastizales, por lo tanto, se incrementaría la infiltración del agua superficial (Lopes et al., 2018; Lozano‑Trejo et al., 2020).
La infiltración de agua en el suelo también puede ser influenciada por la estructura del suelo, una mejora en la calidad de la estructura del suelo promueve un buen drenaje y pue‑ de reducir significativamente la escorrentía superficial y los procesos erosivos (Brasil et al., 2021); favoreciendo de esta manera a la infiltración, lo cual concuerda con nuestros resultados, ya que los bosques presentaron valores superiores, en materia orgánica y capacidad de intercambio catiónico en el suelo que los pastizales. Estudios experimentales previos han demostrado que la capacidad de infiltración y permeabilidad de los suelos están directamente relacionados con la porosidad del suelo (Santos et al., 2021; Zhang et al., 2021), densidad de raíces (Luo et al., 2021) y el crecimiento y descomposición de raíces (Luo et al., 2014; Bork et al., 2021).
El modelo potencial fue el mejor modelo ajustado para el comportamiento de la infiltración del agua en el suelo de los bosques primarios, similares resultados fueron reportados por estudios previos (Karlin et al., 2019; Lozano‑Trejo et al., 2020). Además, las diferencias en el comportamiento de las curvas de infiltración según el tipo de cobertura también fueron reportados por estudios previos (Lozano‑Trejo et al., 2020; Tapia et al., 2020), y esto podría deberse a las diferentes condiciones edáficas y florísticas (Wang et al., 2019; Luna et al., 2020) como en los bosques y pastizales evaluados. Una mejor calidad en las características del suelo (mayor materia orgánica y capacidad de intercambio catiónico) estaría generando una mayor infiltración en los bosques que en los pastizales (Brasil et al., 2021).
Nuestros resultados, de nivel exploratorio, resaltan la gran contribución de los bosques en la conservación del agua de las precipitaciones, en comparación con los pastizales. De esta manera, son importantes las acciones y programas de educación ambiental con la población local, fomento de la recuperación de la cobertura vegetal (sistemas agroforestales, plantaciones, etc.) y gestión sostenible de los recursos forestales maderables y no maderables. Debido a que estas actividades pueden minimizar los impactos ambientales debido a las inundaciones y erosión de los suelos (Lopes et al., 2018).
Desde esta perspectiva sugerimos iniciativas para la gestión y restauración de los pastizales y bosques secundarios para favorecer la conservación de la biodiversidad y el mantenimiento de los servicios ecosistémicos, como la infiltración. Además, en la Amazonia de Madre de Dios son necesarios más estudios para investigar los efectos de la vegetación en la infiltración de agua en el suelo. En futuros estudios se recomienda considerar a más tipos de cobertura vegetal, incrementar el número de muestras, correlacionar la capacidad de infiltración con las características fisicoquímicas del suelo, y considerar a más características del suelo, como por ejemplo, la densidad aparente del suelo.
CONCLUSIONES
Nuestros resultados sugieren que en los bosques primarios, la capacidad de infiltración de agua en el suelo es superior a los pastizales. Además, encontramos que los sitios con mayor abundancia, riqueza y diversidad en la vegetación arbórea tienden a tener una mayor capacidad de infiltración en el suelo. Esto tendría impactos significativos en el comportamiento hidrológico de los ecosistemas tropicales, debido a la transformación de bosques a pastizales generados por las actividades antrópicas.

